


Extraembryonic mesoderm in primate and rodent embryos and models
 0
0 Abstract
Extraembryonic mesenchymal cells/mesoderm cells (Exmes/EXMC) are mesenchymal-like populations located outside the embryo proper during early amniote development. They line the amnion and yolk sac, form the placental villous core, and build the connecting stalk. They are essential for extraembryonic tissue support, early hematopoiesis and vascular development. Yet their developmental origins and lineage relationships remain partially resolved, particularly across species. In rodents, EXMC arise during gastrulation from newly specified mesoderm emerging from the primitive streak. In primates including humans, by contrast, extraembryonic mesenchymal cells (Exmes) are detectable in the post-implantation but pre-gastrulation conceptus, before the formation of gastrulation-derived mesoderm. Here, focusing on the peri-implantation to early-organogenesis time window (CS3-CS9 in primates; E4.5-E8.5 in rodents), we review histological, genetic, and single-cell multi-omic evidence on the emergence, diversification, and functional specialization of pre-gastrulation Exmes/post-gastrulation EXMC, with particular attention to their roles in yolk sac and placental hematopoiesis, vascularization, and tissue homeostasis. We further discuss insights from non-human primate models and human stem cell-based embryo models that generate Exmes/EXMC-like populations in vitro. Finally, we outline key challenges for resolving the origins of these cells, defining conserved and species-specific regulatory programs, and leveraging Exmes/EXMC biology to understand pregnancy loss, placental disorders, early hematopoietic and vascular development.
Keywords
INTRODUCTION
Extraembryonic mesenchymal cells/mesoderm cells (Exmes/EXMC) play a central role in orchestrating early mammalian development. By providing structural support, vascular niches and signalling environments, they enable embryonic growth, extraembryonic hematopoietic and vascular development[1]. In mice, the developmental origin of extraembryonic mesoderm is well defined: it arises from epiblast-derived mesoderm that emerges through the primitive streak during gastrulation and subsequently expands into extraembryonic territories[2]. This model has therefore served as the canonical framework for understanding EXMC formation. However, studies in primates, including humans and non-human primates, have revealed a distinct developmental pattern. Mesenchymal-like cells are already present at post-implantation stages within the chorionic cavity before the onset of gastrulation[3,4]. These early-appearing cells, here referred to as Exmes, occupy the chorionic cavity, surround the yolk sac and amnion, and contribute to the earliest formation of placental villous core. Their presence before gastrulation challenges the classical view that extraembryonic mesoderm is exclusively derived from gastrulation-derived mesoderm.
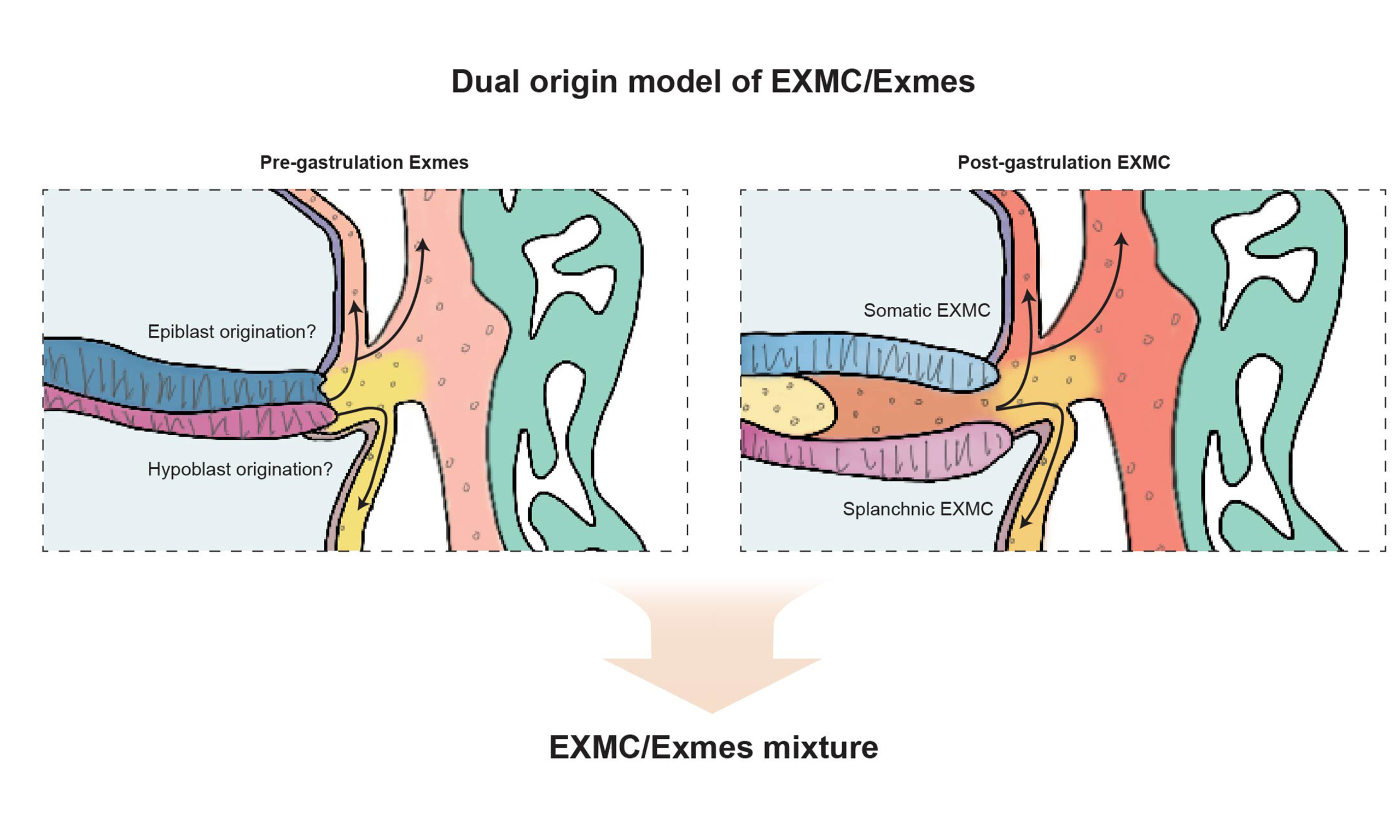
This temporal discrepancy raises a fundamental question: what is the developmental origin of Exmes in primates? Historically, three main hypotheses have been proposed for the origin of primate Exmes, suggesting derivation from trophoblast, epiblast or hypoblast before gastrulation. Early histological studies supported a trophoblastic origin. However, accumulating molecular and lineage tracing evidence has largely challenged this model and instead points to an inner cell mass (ICM)-related origin. Whether Exmes arise directly from the epiblast, from the hypoblast, or from both lineages therefore remains unresolved. Recent advances in single-cell transcriptomics[5,6], spatial profiling[7,8] and stem cell-based embryo models[9-12] have reshaped our understanding of these early primate Exmes lineages[13]. Emerging evidence suggests that primate Exmes are not a homogeneous lineage. In this framework, early Exmes emerge before gastrulation and show molecular features consistent with a hypoblast-related origin. By contrast, later EXMCs arise from epiblast-derived mesoderm during gastrulation and subsequently integrate into extraembryonic tissues. We use Exmes to denote pre-gastrulation mesenchymal-like cells in the primate extraembryonic compartment, and EXMCs to denote gastrulation-derived extraembryonic mesodermal populations homologous to those described in rodents. We retain Exmes/EXMC as an umbrella term for the broader lineage continuum.
In this Review, we revisit the developmental origin, diversification and functions of extraembryonic mesenchymal and mesodermal populations across primates and rodents. We first compare rodent and primate developmental programmes to define this conceptual distinction. We then assess histological, lineage-based and single-cell multi-omic evidence for the origins and transitions of these populations, with particular attention to stem-cell-based embryo models. Finally, we discuss how Exmes/EXMC build the stromal, vascular, signalling and haematopoietic infrastructure of extraembryonic tissues, and how this biology may inform our understanding of placental disorders, pregnancy loss and early blood development.
EARLY EMBRYONIC DEVELOPMENT
Early embryonic development in rodents
Understanding early mouse embryogenesis provides a canonical framework for understanding how extraembryonic lineages-including EXMC-are specified and patterned. By embryonic day 3.5 (E3.5), the mouse embryo has formed a blastocyst consisting of an outer trophectoderm (TE) that encloses the ICM[2]. Around E4.0, the blastocyst hatches from the zona pellucida, and the ICM diversifies into epiblast and primitive endoderm (PrE)[2]. Implantation occurs at approximately E4.5. Unlike primates, in which the polar TE (pTE) mediates attachment to the endometrium[14], rodent implantation is initiated by the mural TE (mTE)[14]. This geometric difference is thought to contribute to the divergent morphologies of the post-implantation embryos: a cup-shaped structure in mice versus bilaminar disc in primates.
Following implantation, the mTE deeply invades the maternal endometrium. PrE differentiates into visceral endoderm (VE), which envelops the basal surface of the epiblast, and parietal endoderm (PE), which spreads along the inner surface of the trophoblast[2] [Figure 1A]. By E5.5, as trophoblast cells proliferate and invade, mTE gives rise to the extraembryonic ectoderm (ExE) and the ectoplacental cone (EPC)[15]. The pro-amniotic cavity originates from a hollowing process rather than apoptosis-driven cavitation. Around E5.5, polarized epiblast cells begin to generate small intercellular luminal spaces through separation of the apical membranes, likely mediated by anti-adhesive molecules such as podocalyxin[16,17] [Figure 1B]. Similarly, the ExE also develops a podocalyxin-coated intermembranous space. As development proceeds, these initially separate luminal structures subsequently fuse to form a single mature pro-amniotic cavity spanning both the EPI and ExE[16,17] [Figure 1C]. During this window, the primary body axes are progressively established. Using the epiblast as a reference, the side closest to the ExE is designated proximal and the opposite side distal. A specialized population of VE, the distal visceral endoderm (DVE), emerges at the distal tip of the conceptus[18] [Figure 1B].
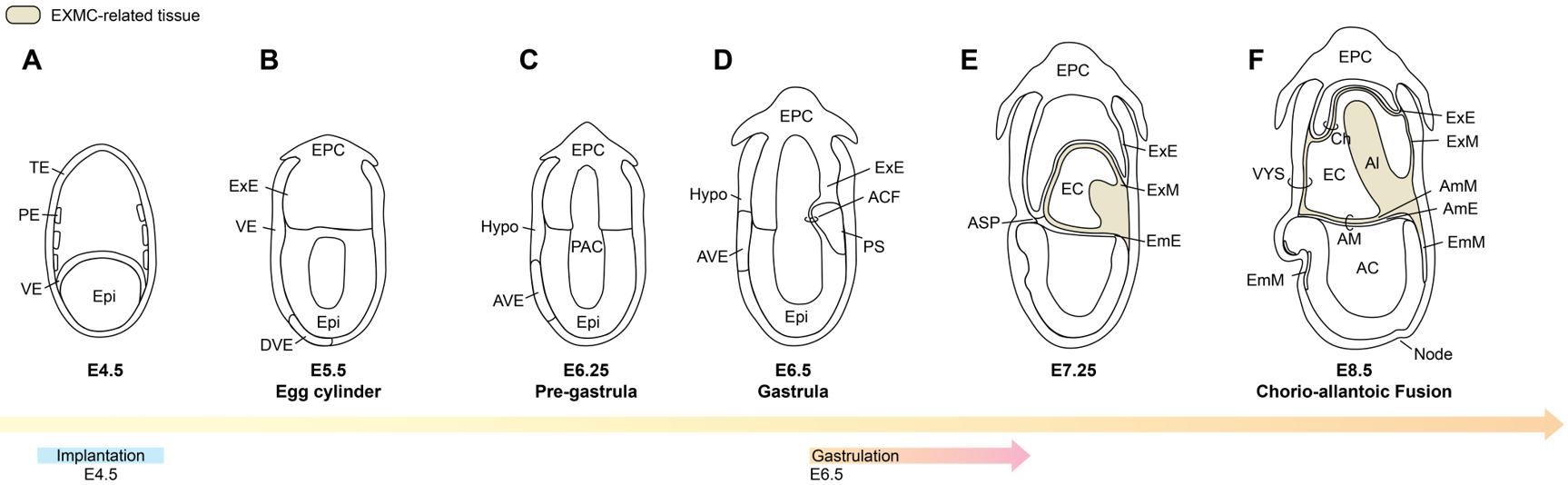
Figure 1. Developmental patterns from the implantation stage (A) to gastrulation (D), and to the early post-gastrulation phase (E and F) in rodent embryos. TE: Trophectoderm; PE: parietal endoderm; VE: visceral endoderm; Epi: epiblast; Hypo: hypoblast; EPC: ectoplacental cone; DVE: distal visceral endoderm; AVE: anterior visceral endoderm; PAC: pro-amniotic cavity: ACF: amniochorionic fold; PS: primitive streak; ASP: anterior separation point; ExE: extraembryonic ectoderm; ExM: extraembryonic mesoderm; AmE: amniotic ectoderm; AmM: amniotic mesoderm; EmE: embryonic ectoderm; EmM: embryonic mesoderm; AM: amnion; AC: amniotic cavity; VYS: visceral yolk sac; EC: exocoelomic cavity; Ch: chorion; Al: allantois. Image created by Adobe Illustrator.
Around E6.25, DVE cells migrate from the distal position towards the proximal anterior region of the embryo[19]. After migration, these cells act as a signalling centre that patterns the anterior-posterior (A-P) axis[20]. The future head side of the embryo is defined as anterior and the tail side as posterior. Because the migrated DVE settles on the anterior side, this cell population is termed anterior visceral endoderm (AVE)[13,20] [Figure 1C]. Once the A-P polarity is established by AVE, the embryo undergoes gastrulation at approximately E6.5[13,18] [Figure 1D]. A primitive streak forms at the posterior margin of the epiblast, from which newly mesodermal cells emerge[13]. This nascent mesoderm bifurcates into embryonic mesoderm and EXMC, which will populate extraembryonic tissues.
Rapid posterior morphogenesis drives the accumulation of EXMC and ExE, giving rise to the amniochorionic fold (ACF), a structure that contributes to the formation of both amnion and chorion during early embryogenesis[21,22] [Figure 1D]. As the ACF extends, it encloses a cavity that will become the extraembryonic coelom (exocoelom)[22] [Figure 1E]. Amniotic ectoderm (AmE) spreads along this fold towards the anterior separation point (ASP)[21,22], where it meets the ExE at the anterior pole. This configuration partitions the original pro-amniotic cavity into two compartments [Figure 1E]: an amniotic cavity that envelops the embryo proper, and the exocoelom, bounded by ExE and extraembryonic tissues. Subsequent morphogenesis and expansion of extraembryonic tissues contribute to the formation of the visceral yolk sac and separation of the amnion and chorion[21,22].
By E8.5, the allantois, derived from extraembryonic mesoderm, extends through the exocoelom towards the mural side of the visceral yolk sac and fuses with the chorion. This chorionic-allantoic fusion permits extraembryonic mesoderm-derived blood vessels to invade the chorionic trophoblast, establishing the nascent chorioallantoic placenta[2,15,23] [Figure 1F].
Pioneering clonal analysis and transplantation studies by Lawson[24], Beddington[25] and colleagues mapped the lineage relationships between the epiblast and its mesodermal derivatives, demonstrating that cells ingressing through the posterior region of the primitive streak contribute prominently to mouse EXMC. These classical experiments provided the first dynamic view of EXMC generation and dispersal in vivo, and laid the foundation for subsequent molecular and single-cell studies that now revisit EXMC ontogeny with higher resolution.
Early embryonic development in primates
In primates, the developmental stage from fertilization to approximately 8 weeks post-fertilization constitutes the embryonic period. Based on morphology, somite number and crown-rump length, this interval is subdivided into 23 Carnegie stages (CS)[26,27]. At CS3 (around days 4-5 post-fertilization), the first lineage segregation produces the TE, which will subsequently contribute to the trophoblastic shell and placental villi, and the ICM, which gives rise to the embryo proper and major extraembryonic lineages, including the amnion and yolk sac[28]. A second fate specification event then occurs within the ICM[28], in which pluripotent epiblast cells, marked by POU5F1 and NANOG, intermingle with hypoblast precursors expressing GATA4/6 in a characteristic salt-and-pepper pattern[29], anticipating segregation into epiblast and hypoblast layers [Figure 2A].
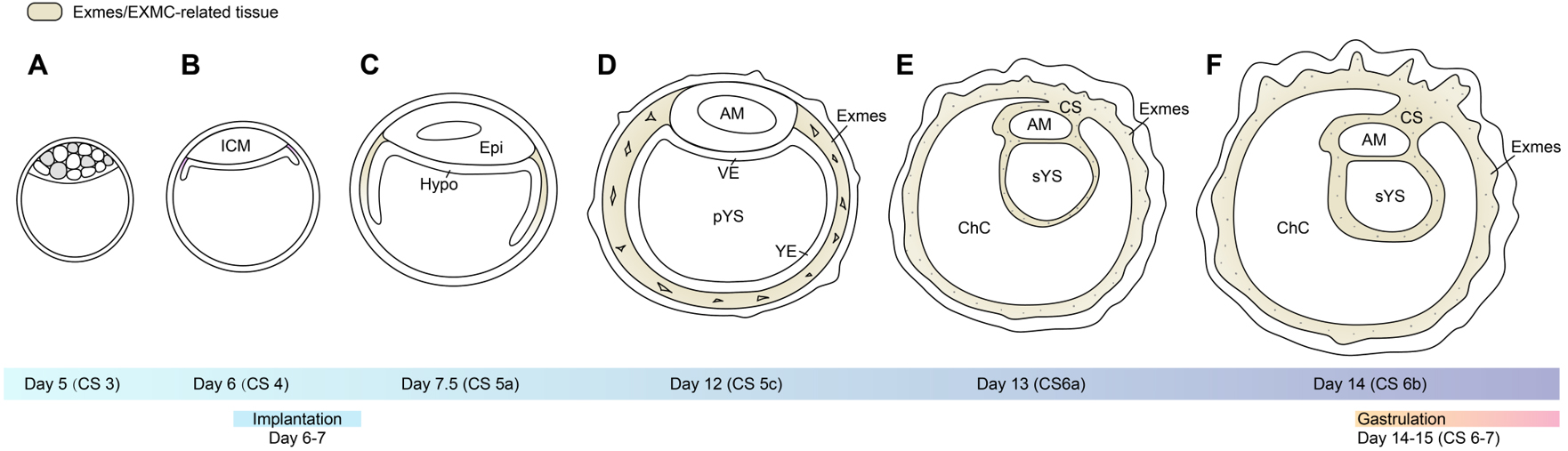
Figure 2. Developmental patterns and cell lineage differentiation from the blastocyst stage (A) through implantation (B) to pre-gastrulation (F) in primate embryos. ICM: Inner cell mass; Epi: epiblast; Hypo: hypoblast; AM: Amnion; VE: visceral endoderm; YE: Yolk sac endoderm; pYS: primary yolk sac; Exmes: extraembryonic mesenchymal cells; sYS: secondary yolk sac; CS: connecting stalk; ChC: chorionic cavity. Image created by Adobe Illustrator.
By CS4, following hatching from the zona pellucida, the blastocyst initiates implantation into the endometrium[30], at which point the epiblast and hypoblast have largely resolved into their respective spatial domains [Figure 2B]. At CS5, the primate conceptus becomes deeply embedded within the endometrium (fully interstitial in humans and more superficially in many non-human primates[30]) [Figure 2C]. pTE proliferates and invades the maternal tissue, and through cell fusion and/or endoreplication or endomitosis gives rise to a specialized multinucleated syncytial layer, often referred to as the primitive syncytiotrophoblast (pSTB)[31], which mediates the earliest post-implantation trophoblast invasion[32].
Concurrently, a bilaminar embryonic disc with three extraembryonic cavities is established. The epiblast differentiates into the amniotic epithelium and encloses the amniotic cavity; the hypoblast spreads beneath the epiblast, while yolk sac endoderm extends ventrally and closes to form the primary yolk sac (pYS)[3,4] [Figure 2D]. The interposed space between the yolk sac and the trophoblast delineates the chorionic cavity. At this stage, the chorionic cavity is traversed by a loosely organized meshwork of mesenchymal cells
At the maternal-fetal interface, the primary villi are composed of primitive cytotrophoblast (pCTB) and pSTB[33] [Figure 3A and B]. At CS6, the chorionic cavity expands as trophoblast cells invade the decidual tissue, forming chorionic villi filled with a mesenchymal core[34], known as secondary villi[33] [Figure 3C and D].
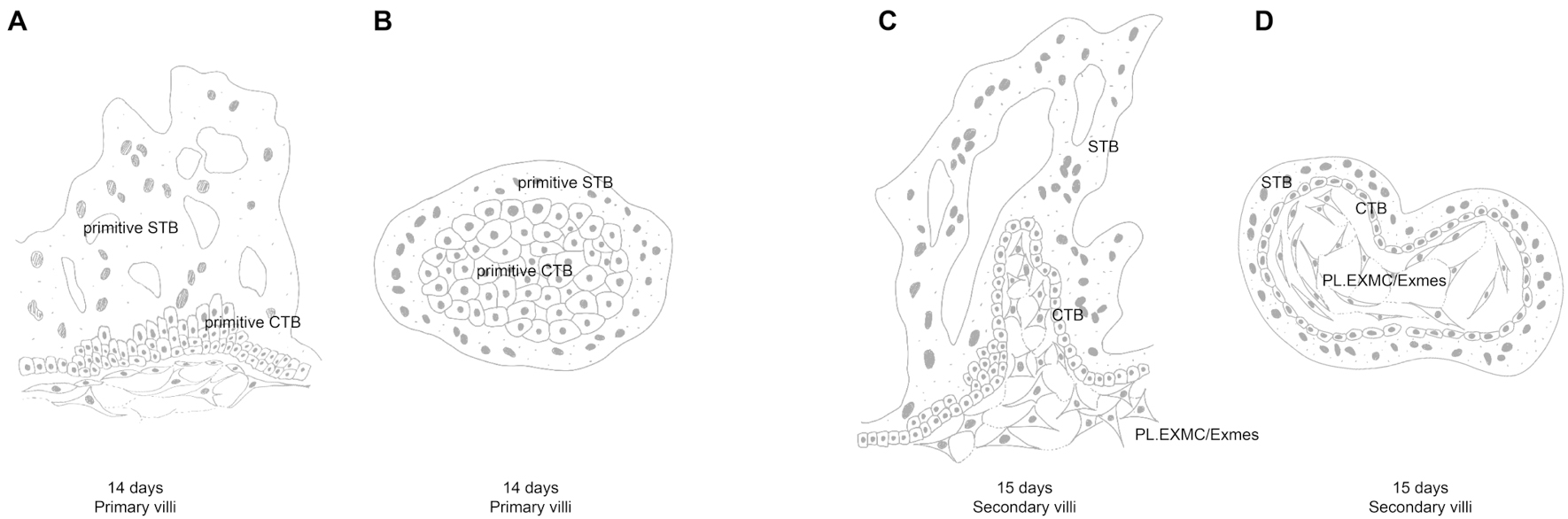
Figure 3. pSTB and pCTB invade the uterus to form primary villi (A and B). Exmes subsequently infiltrates these primary villi, forming secondary villi with a mesenchymal core (C and D). Image created by Adobe Illustrator.
At CS6, as the bilaminar embryonic disc expands, the Exmes form a continuous mesenchymal sheet that envelops the outer surface of the amnion and yolk sac, as well as the inner surface of the trophoblast. Within this mesenchymal layer, multiple small vacuoles begin to emerge. Shortly thereafter, vacuoles within the chorionic cavity progressively enlarge, coalesce, and reorganize into a single dominant extraembryonic cavity, resulting in the persistence of the connecting stalk as the major residual structure[35] [Figure 2E and F]. This structure anchors the embryo to the chorionic trophoblast and marks the posterior pole of the disc, with the opposite side corresponding to the future anterior region. Around CS6a, peripheral expansion and remodelling of Exmes within the chorionic cavity drive a characteristic transformation of the yolk sac. The primary yolk sac is gradually compressed and collapses[4] [Figures 2D and 4A and B]; a portion of its lumen becomes sequestered on the mural side of the chorionic cavity, whereas the residual structure remodels into the secondary yolk sac (sYS)[4] [Figure 2E and 4C]. Thus, Exmes dynamics are intimately coupled to yolk sac morphogenesis and the establishment of A-P polarity.
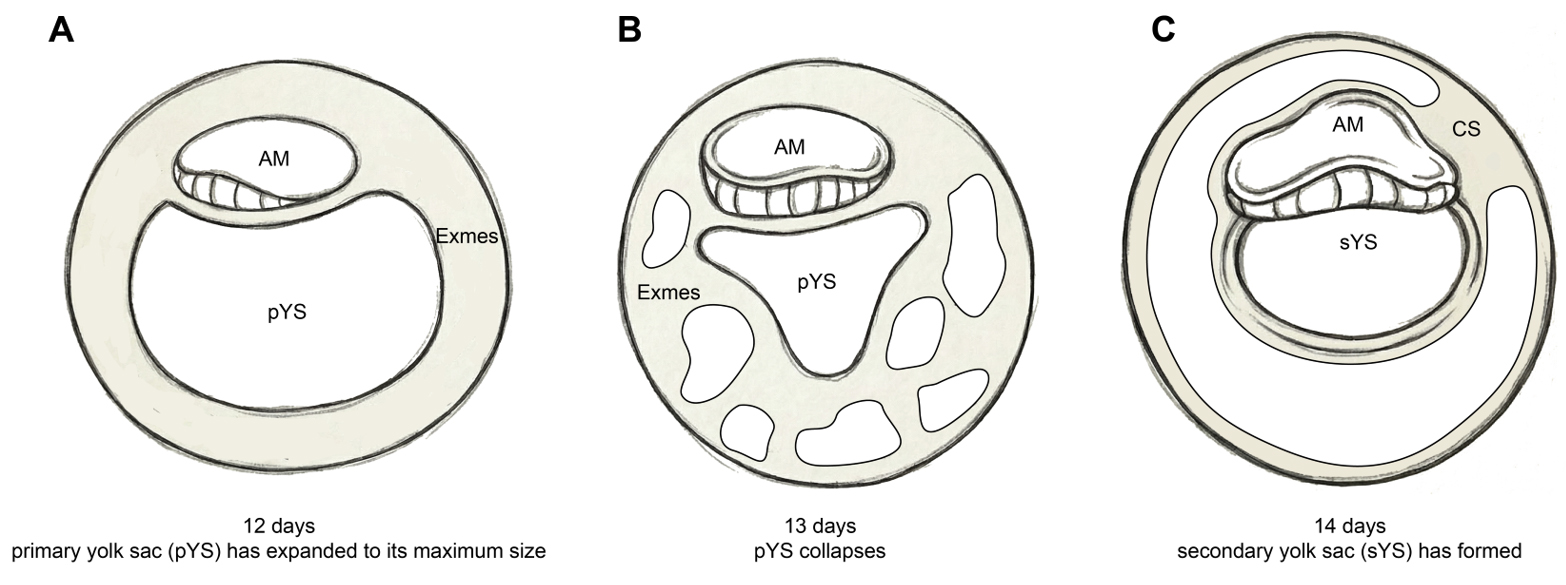
Figure 4. Developmental schema of the primate embryonic yolk sac. The primary yolk sac (A) undergoes progressive collapse and remodeling (B), giving rise to the secondary yolk sac (C). The extraembryonic space is populated by Exmes prior to gastrulation. AM: Amnion; pYS: primary yolk sac; sYS: secondary yolk sac; Exmes: extraembryonic mesenchymal cells. Image created by Adobe Illustrator.
By CS6b, a primitive streak anlage appears at the caudal (posterior) margin of the epiblast, signalling the onset of gastrulation[3,5]. The primitive node extends cranially from the streak beneath the overlying ectoderm and initiates the notochordal process [Figure 5A], which will give rise to the axial mesoderm. At CS7, epiblast cells ingress through the primitive streak and migrate ventrally, interposing a nascent mesodermal layer between ectoderm and hypoblast[36] [Figure 5B]. Concomitantly, definitive endoderm cells derived from the epiblast displace the hypoblast lining the embryonic region [Figure 5C].
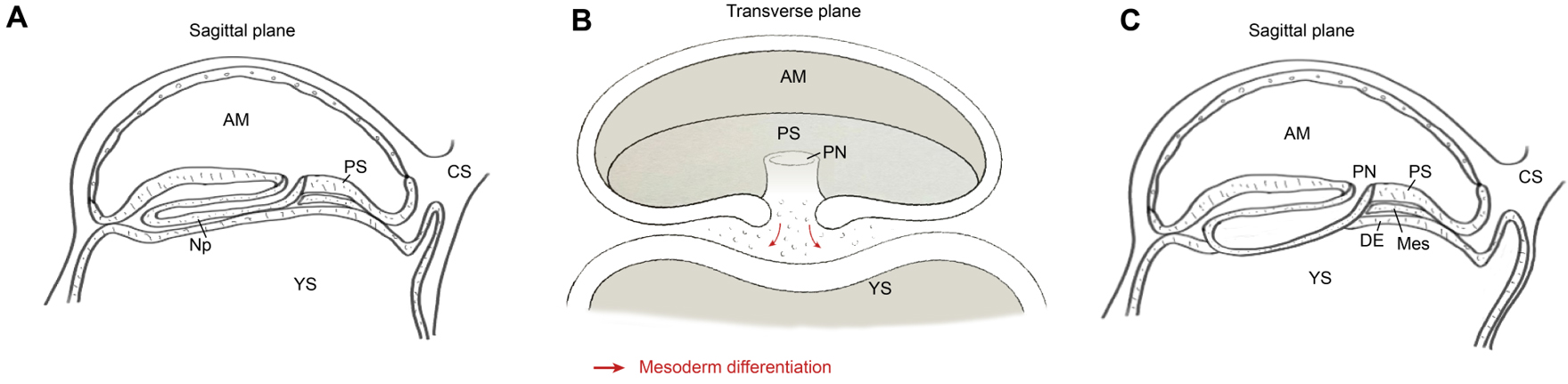
Figure 5. Schematic diagrams in sagittal (A and C) and transverse (B) planes illustrating the generation of nascent mesoderm and definitive endoderm during gastrulation, after which the bilaminar structure transforms into a trilaminar structure. Following gastrulation, the mesoderm migrates outward and differentiates into EXMC. The red arrow indicates that the differentiation of the mesoderm during gastrulation originates from the proliferation and differentiation of the epiblast towards the ventral side. AM: Amnion; PN: primitive node; PS: primitive streak; YS: yolk sac; CS: connecting stalk; Np: notochordal process; DE: definitive endoderm; Mes: mesoderm. Image created by Adobe Illustrator.
In this framework, early-appearing Exmes in the chorionic cavity coexist with, and are later complemented by, gastrulation-derived extra-embryonic mesoderm, raising fundamental questions about how these temporally and spatially distinct extraembryonic mesodermal populations are related in lineage diversification and function.
At CS8, continued proliferation and morphogenesis of the epiblast drive early folding as well as elongation of the embryonic disc. Gastrulation proceeds through CS9, at which point the three definitive germ layers are established and the blueprint for organogenesis is in place. Mesodermal tissues now flank the notochord bilaterally and diversify into paraxial, intermediate, lateral plate and extraembryonic mesoderm subclusters[36] [Figure 6]. Paraxial mesoderm segments to form somites[37] adjacent to the closing neural groove. The lateral plate mesoderm, which remains continuous with the extraembryonic mesoderm, splits with the emergence of the intraembryonic coelom into somatic and splanchnic layers that will contribute to the body wall and the circulatory system[38]; the anterior lateral plate mesoderm initiates cardiac morphogenesis. Within the extraembryonic mesoderm of the yolk sac, blood islands containing haemato-endothelial progenitors progressively expand[39], foreshadowing subsequent vascularization of both embryonic and placental territories.
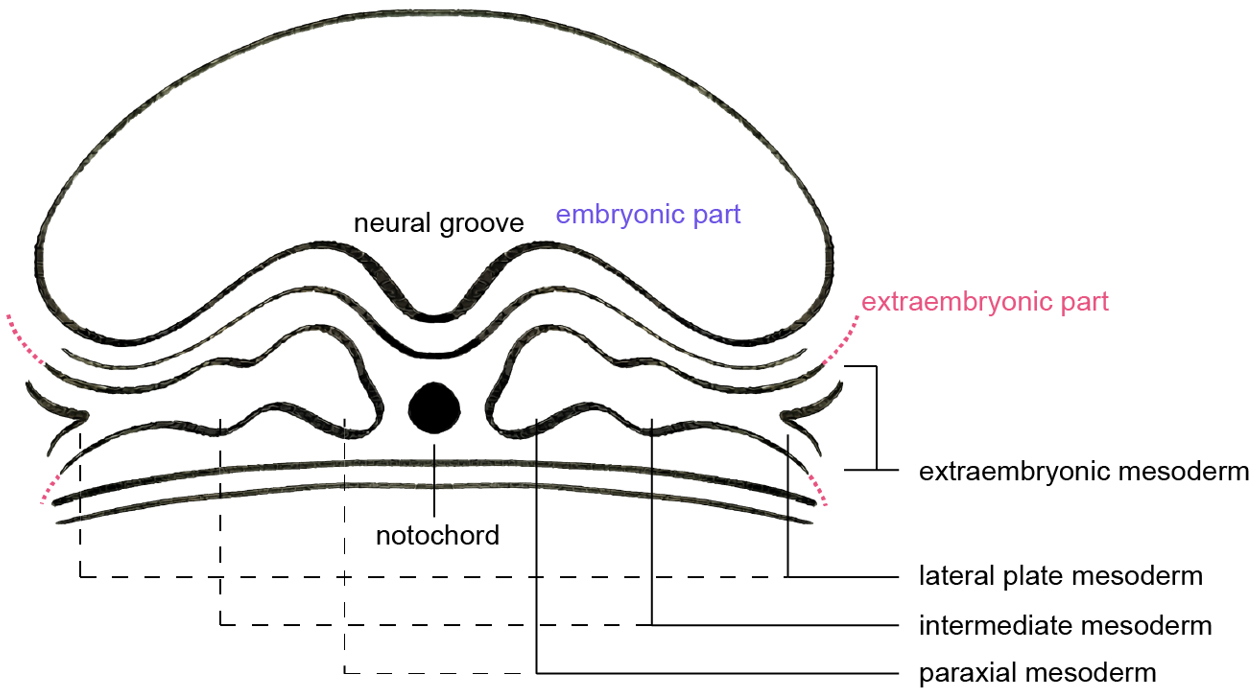
Figure 6. Schematic of a transverse plane through the post-gastrulation three-germ-layer structure, showing the mesoderm differentiating into paraxial, intermediate, lateral plate, and extraembryonic mesoderm. Image created by Adobe Illustrator.
Non-human primate embryos broadly recapitulate the developmental programme of human embryos and therefore provide indispensable models for studying primate embryogenesis. Nonetheless, important species-specific differences exist. A key distinction lies in implantation mode: human embryos undergo complete interstitial implantation within the endometrium[40], whereas many non-human primates exhibit a more superficial, primitive implantation[30].
Morphologically, human blastocysts at embryonic days 10-11 display a primary yolk sac that is partially filled by a loose reticular meshwork of extraembryonic tissue. By contrast, comparable extraembryonic structures have not been reported at similar stages in macaques (Heuser, Streeter), baboons (Hendrickx, Houston) or rhesus monkeys[3]. These disparities may reflect differences in yolk sac morphogenesis and extraembryonic patterning that are secondary to distinct implantation strategies.
In primates, the first descriptions of extraembryonic mesoderm arose from careful histological analyses of human and macaque conceptuses by Streeter, Hertig, Heuser, Wislocki and others. Streeter’s classic account of an approximately 11-day human embryo, the "Miller ovum" (1926)[41], noted that the embryo and primary yolk sac were enveloped by a reticular primitive mesoblast[41] (extraembryonic mesoderm) that contributed to the yolk sac, early placenta and allantois. Because a primitive streak was not yet evident at this stage, Streeter proposed that this primitive mesoderm might derive from adjacent multipotent trophoblast. Subsequent work, however, has challenged this trophoblastic origin hypothesis, instead raising the possibility of contributions from hypoblast and/or epiblast lineages, and highlighting the need for modern lineage tracing and single-cell approaches to revisit the ontogeny of primate Exmes/EXMC.
Exmes BEFORE GASTRULATION IN PRIMATES
From the perspective of embryonic-disc architecture, rodents and primates adopt fundamentally different strategies that shape the extent and organization of extraembryonic territories. In the mouse, the cup-shaped embryonic disc fills most of the blastocyst cavity, with the pro-amniotic cavity and ExE occupying the remaining space. This compact arrangement is often linked to the relatively short gestation and streamlined extraembryonic morphology. By contrast, primate development is characterized by more expansive cavities and elaborated extraembryonic tissues. The embryo proper remains as a flat, bilaminar disc of epiblast and hypoblast floating within a large blastocyst cavity. This broad extraembryonic compartment provides the structural context in which multiple extraembryonic lineages, including Exmes, can emerge and diversify before the onset of gastrulation.
In primate embryos, extraembryonic mesenchyme appears in close temporal association with primary yolk sac formation. At CS5, yolk sac endoderm proliferates and spreads along the inner surface of the trophoblast, progressively lining and enclosing the blastocyst cavity. Completion of this epithelial sheet generates a ring-like structure that defines the primary yolk sac. The inner layer of the primary yolk sac is composed of extraembryonic endoderm (yolk sac endoderm), whereas the space between its outer surface and the overlying trophoblast is filled by a loose, reticulated meshwork of extraembryonic mesenchyme [Figure 2D]. These cells constitute the earliest morphologically recognizable Exmes population in primates and will subsequently remodel into the outline of the yolk sac, amnion and stromal cores of nascent placental villi.
THE INVASION AND CONTRIBUTION OF Exmes/EXMC TO EXTRAEMBRYONIC TISSUES
Exmes constitute the mesenchymal component of extraembryonic organs, including the amnion, yolk sac and inner core of the placenta. They populate and structurally support adjacent epithelial or epithelial-like layers derived from distinct lineages: epiblast-derived amnion, hypoblast-derived yolk sac endoderm, and trophectoderm-derived placental villi. The early connecting stalk, which will give rise to the umbilical cord, is also largely composed of Exmes, highlighting the central role of these cells in physically linking the embryo to the extraembryonic compartment and maternal tissues.
In rodents
In rodent embryos, EXMC arise via gastrulation and, by around E7.5, contribute extensively to the visceral yolk sac, which occupies the space between the amnion and chorion. The molecular control of rodent extraembryonic mesoderm development has been dissected in considerable detail. Gene-targeting studies have revealed that loss of BMP2 or BMP4, as well as mutations in BMP receptors, causes severe defects in extraembryonic tissues and frequently result in early embryonic lethality[42], underscoring the dependence of EXMC formation and function on BMP signalling.
For the amnion
On the amniotic side, EXMC contribute to the formation of the mesodermal layer that overlays the amniotic epithelium. Cells in this amniotic mesoderm exhibit high expression of mesoderm-associated genes such as Fn1 and Foxf1, consistent with their mesenchymal identity, and reduced expression of epithelial markers including Epcam and Cldn7. In addition, amniotic mesoderm cells show robust expression of Postn, Bmp2 and Bmp4, among other factors[21]. Functional studies indicate that BMP2 is required for normal morphogenesis of the amnion and chorion: BMP2 deficiency leads to abnormal closure of the pro-amniotic canal and subsequent defects in amniotic and chorionic development[43].
For the yolk sac
Within the visceral yolk sac, local thickening of EXMC is observed around E7.5, coincident with the emergence of blood islands along the lateral walls. These structures comprise hemato-endothelial progenitors embedded in an EXMC-derived stromal matrix and represent the earliest site of primitive hematopoiesis. Transforming growth factor-β (TGF-β) signalling is critical for maintaining both the angiogenic and primitive hematopoietic functions of EXMC in the yolk sac[44]. TGF-β promotes EXMC-derived angiogenesis by enhancing endothelial cell formation and maintaining vascular stability[45,46]. Homozygous mutation of the basic helix-loop-helix transcription factor Hand1 results in embryonic lethality associated with defective EXMC development, leading to yolk sac malformations and abnormal cardiac looping[47]. These findings position EXMC at the nexus of extraembryonic vascular, hematopoietic and cardiac development. They further demonstrate that perturbations of extraembryonic mesoderm propagate across multiple embryonic and extraembryonic organ systems.
For the chorio-allantoic fusion and placenta
On the placental side, EXMC together with ExE form the chorionic plate. By E8.5, the allantois—entirely derived from extraembryonic mesoderm—extends through the exocoelom towards the mural side and fuses with the chorion[48,49] [Figure 1F]. Successful chorio-allantoic fusion permits blood vessels originating in the extraembryonic mesoderm of the allantois to invade the chorionic trophoblast. The intimate interdigitation of these fetal vessels with maternal blood sinuses establishes the labyrinthine layer of the mouse placenta, which is indispensable for effective maternal-fetal gas and nutrient exchange and therefore for embryonic survival and normal growth[15]. Failure of chorio-allantoic fusion results in placental insufficiency and embryonic lethality.
BMP4 plays a pivotal role in allantoic development: at E6.5, Bmp4 is highly expressed at the posterior end of the primitive streak, and by E7.5 it is strongly upregulated in the emerging allantois, where it promotes mesodermal differentiation and outgrowth[50]. Disruption of the adhesion molecule VCAM-1 or its receptor integrin α4, as well as combined deficiencies in BMP5 and BMP7, impairs or abolishes chorio-allantoic fusion[42,51]. Moreover, Rspo3 deficiency prevents vascular invasion of the chorion[52], while loss of Hgf or Wnt2 perturbs chorionic branching morphogenesis and labyrinth formation[51]. Together, these studies highlight a complex signalling network in which BMP, WNT, RSPO and growth factor pathways orchestrate EXMC differentiation, allantoic elongation and placental morphogenesis.
In primates
For the amnion
During CS5a, continued thickening and cavitation of the epiblast generates the pro-amniotic cavity[53,54]. By CS5c, as Exmes progressively fill the extraembryonic space, the amnion is organized as a bilayered structure comprising an epithelial sheet and an overlying Exmes-rich mesenchyme. In this configuration, Exmes provide a stromal scaffold that anchors the embryo in a relatively fixed position within the chorionic cavity. Subsequent asymmetrical growth and patterning along the A-P axis cause regional stretching of the Exmes-rich domain towards the posterior pole, where it condenses into the connecting stalk[35]. This early stalk not only serves as a mechanical tether but also demarcates the posterior side of the bilaminar disc, coupling Exmes morphogenesis to axial specification.
For the yolk sac
Following primary yolk sac formation, a mesenchymal cell population emerges overlying the outer surface of the yolk sac endoderm, recognized as yolk sac Exmes, which provide structural and signaling support for subsequent primitive haematopoiesis. From around CS6b, focal aggregations of Exmes become evident along the yolk sac, corresponding to the earliest rudiments of blood islands, marking the onset of primitive wave of yolk sac hematopoiesis. By CS7, endothelial cell-lined blood channels emerge, and by CS8, primitive hematopoietic cells are observed within these nascent vessels[4], marking the onset of the pro-definitive wave of yolk sac hematopoiesis. Recent human yolk sac cell-atlas studies have further shown that the yolk sac endoderm provides key hematopoietic growth factors that support the hemato-endothelial differentiation of Exmes/EXMC-derived progenitors[55], underscoring a functional crosstalk between endodermal and mesodermal compartments.
For the placenta
Around 13-15 days post-fertilization[34], Exmes/EXMC invade the solid primary villi formed by pCTB and pSTB, generating the mesenchymal core of the nascent placental villi. This Exmes/EXMC -derived villous stroma progressively differentiates into vascular endothelium, perivascular mesenchyme and placental macrophages-Hofbauer cells (HBCs). HBCs, in turn, contribute to vascular remodelling and angiogenic signalling. Notably, angiogenic activity and the emergence of embryonic macrophages precede the establishment of a fully functional fetoplacental circulation, raising the possibility that the placenta itself may act as an early extraembryonic haematopoietic niche. Consistent with prevailing models, HBCs may arise either from yolk sac-derived progenitors that migrate to the placenta or from hemogenic endothelium within placental vasculature. However, the precise developmental origin and relative contribution of these sources to primitive haematopoietic lineages in the placenta remain unresolved and will be discussed in subsequent sections.
In summary, the brief but highly dynamic window (CS3-CS9 in primates; E4.5-E8.5 in rodents) from implantation to gastrulation lays the groundwork for healthy embryogenesis. Proper early patterning and maturation of extraembryonic tissues-anchored structurally and functionally by Exmes/EXMC-are essential for sustained intrauterine growth and a physiologically normal pregnancy. The amnion safeguards the embryonic milieu, the yolk sac supports primary hematopoiesis, the allantois enables gas and nutrient transport, and the placenta orchestrates maternal-fetal exchange and immune regulation[3,15,56-58]. These processes must unfold rapidly and with exquisite spatiotemporal precision, with Exmes/EXMC undergoing parallel maturation to build the stromal, vascular and immunological infrastructure of extraembryonic organs. Yet our understanding of these events remains fragmentary.
Ethical constraints[59,60] and the scarcity of very early human and non-human primate samples present major obstacles, and species-specific variation further complicates comparisons. In rodents, extraembryonic mesoderm can be clearly traced to mesoderm emerging through the primitive streak during gastrulation. By contrast, in primates, as in other eutherian mammals, mesenchymal-like cell types in the extraembryonic compartment are observed before the onset of gastrulation[61,62]. As a result, three principal lineage candidates have been proposed for the earliest Exmes: trophoblast, epiblast or hypoblast. At present, there is no definitive consensus on the developmental origin, lineage relationships or differentiation trajectories of Exmes in very early primate pregnancy, making this a central open question that next-generation in vivo and in vitro models must address.
DEVELOPMENTAL ORIGINS OF Exmes IN PRIMATE EMBRYOS
Because the experimental inaccessibility of the post-implantation to pre-gastrulation window and the marked species divergence in early extraembryonic lineages, the developmental origin of Exmes in primate embryos has long remained obscure. Recent maturation of single-cell and spatial multi-omics technologies, together with advances in stem-cell-based embryo models and organoid systems, longstanding assumptions about Exmes ontogeny are now being revisited with increasing precision.
The hypothesis that Exmes originate from the trophoblast lineage
Historically, the origin of Exmes in primates has been deeply debated. Early embryologists-most notably Heuser and Streeter-proposed that trophoblast cells possess unexpected multipotency and could differentiate into Exmes, blood vessels and even amniotic epithelium. This proposal was based on serial histological examinations of rare human and macaque specimens, most famously exemplified by Streeter’s description of an approximately 11-day human conceptus ("Miller ovum")[41], in which a reticular mesenchymal network filled the blastocyst cavity and formed a thin, one- to two-cell layer lining the inner surface of the trophoblast[63]. Because a primitive streak had not yet formed in these embryos, the close physical association between the trophoblast and Exmes, together with the apparent in situ emergence of villous mesenchyme, led Hertig and others to propose a trophoblastic origin for these cells.
However, this hypothesis was derived from static observations of conceptuses at differing stages and lacked mechanistic or lineage-based evidence. Subsequent higher-resolution morphological and molecular analyses have challenged the trophoblast-origin model. For example, although epithelial-to-mesenchymal transition (EMT) can induce mesenchymal marker expression (including vimentin, fibronectin and N-cadherin) in MAP3K4-deficient mouse trophoblast stem cells, there is no compelling in vivo evidence to support EMT-mediated mesenchymal differentiation of trophoblast cells within human villi[64]. Furthermore, detailed histology of zona-free rhesus blastocysts by Enders and colleagues demonstrated a continuous basal lamina beneath pCTB that physically separates trophoblast from Exmes and prevents pCTB detachment into the extraembryonic mesenchyme[65]. A similar basal lamina was observed in approximately 10-day human embryos[66]. These observations strongly argue against direct trophoblast-to-mesenchyme conversion. Finally, expression of ICM-specific transcription factors (SOX2, OCT4, REX1, NANOG) within villous mesenchyme[67] further contradicts a trophoblastic origin and instead indicates a lineage relationship to the ICM. Collectively, these datasets have largely disfavoured the trophoblast-origin hypothesis.
The hypothesis that Exmes originate from the ICM
Accumulating evidence-morphological, molecular and transcriptomic-favours derivation of Exmes from the ICM, although the relative contributions of epiblast versus hypoblast remain unresolved.
Epiblast as a potential source of Exmes
An epiblast origin of primate Exmes has long been considered plausible because rodent EXMC are unequivocally derived from epiblast cells ingressing through the primitive streak during gastrulation. Morphological evidence supports this view. Re-analysis of human and rhesus monkey Carnegie embryos by Luckett[3] revealed that primate Exmes development is homologous to that of other eutherian mammals, contradicting Hertig’s hypothesis that trophoblast cells are pluripotent and generate Exmes. Earlier, Florian had inferred from a 12-13-day human embryo that at least a portion of Exmes originate from the caudal epiblast margin[68]. Luckett expanded this idea by proposing that this caudal epiblast represents a precociously forming primitive streak at CS5c-CS6 and may serve as the source of all Exmes in primates[3]. Additionally, persistent expression of pluripotency-associated genes (OCT4, NANOG) in placental mesenchyme[67] supports a potential epiblast lineage relationship.
Molecular evidences from in vitro cultured cynomolgus embryos provide further support. Single-cell analyses identified epiblast-derived Exmes-like cells expressing MIXL1, MESP1, ETS1, PODXL and EPHA4 by day 10. By day 14, cells expressing TBXT, MIXL1 and MESP1 emerged, marking gastrulation onset. A parallel population of Exmes (expressing PODXL, ETS1, PITX1, LUM, NID2) was detected at day 12; by day 14, these cells expressed PITX1, FOXF1, PDGFRA and HAND1 but not TBXT, MIXL1 or MESP1[69]. These findings raise the possibility that epiblast cells may contribute to Exmes even before overt primitive streak formation. Therefore, an epiblast-derived contribution to Exmes prior to gastrulation cannot yet be excluded. Resolution of this question will require future synthetic embryology platforms that leverage lineage-restricted gene editors and high-fidelity barcoding.
Hypoblast as a potential source of Exmes
An alternative model posits that Exmes might arise from before gastrulation. In early primate embryos, Exmes often appear morphologically continuous with extraembryonic endodermal cells lining the primary yolk sac. This morphological continuity supports a hypoblast origin. Historical anatomical studies provide initial evidence for this model. Gerard (1932)[70], studying pre-primitive-streak chimpanzee embryos, proposed that the reticular extraembryonic mesenchyme derives from hypoblast. Enders later reported that visceral and yolk sac endoderm can delaminate and insert into the space between the primary yolk sac and the primitive pCTB, potentially giving rise to Exmes. Ultrastructural studies further revealed free or loosely attached transitional cells in this region that contain dense endoplasmic reticulum bodies characteristic of later Exmes maturation[58]. More compelling molecular data come from early post-implantation cynomolgus embryos. In these specimens, early Exmes expressed hypoblast markers GATA4 and GATA6, whereas primitive streak markers (TBXT, MIXL1, CDX2) were diminished. These expression patterns suggest a hypoblast identity rather than epiblast or streak origin[71]. In vitro embryo models reinforce this interpretation. In human ESC-derived and cynomolgus embryo-like structures, PrE cells and Exmes share markers including PDGFRA, GATA4, GATA6 and NID2[72,73]. Moreover, BMP4-based induction of PrE produces Exmes-like cells (LUM+, ANXA1+, LIX1−, TBXT−)[61], consistent with a hypoblast-origin pathway.
Dual origin of Exmes/EXMC with recent research
Classical embryological studies by Luckett[3], Enders[58,65], King[58] and others collectively challenged the idea that early Exmes arise from a pluripotent trophoblast lineage. Instead, they supported an origin within the ICM, via either epiblast or hypoblast intermediates. Notably, however, these early models did not distinguish between pre-gastrulation Exmes and post-gastrulation EXMC, and largely treated them as a single, continuous population. Recent work has begun to resolve this conceptual limitation by distinguishing early Exmes from later EXMC as temporally and molecularly distinct, yet developmentally connected, phases within a broader continuum. Under this revised framework, Exmes/EXMC in primates are increasingly understood to arise from two ontogenetically and temporally distinct sources.
Prior to gastrulation, the extraembryonic compartment is populated by mesenchymal cells whose transcriptional profiles closely resemble those of the hypoblast[72,73]. These early Exmes are thought to originate from hypoblast-derived cells that delaminate and migrate into the space between the primary yolk sac and the trophoblast. By contrast, following the onset of gastrulation, epiblast-derived mesoderm emerges through the primitive streak and expands into extraembryonic territories, in a manner analogous to that observed in rodents. This second wave gives rise to EXMC, which subsequently intermix with pre-existing Exmes to form a composite mesenchymal population within extraembryonic tissues.
Owing to ethical constraints and the limited availability of human material, support for this dual-origin model has largely relied on studies in non-human primate embryos and stem cell-based embryo models.
In vivo evidence
In early post-implantation cynomolgus embryos (CS5), Exmes are already transcriptionally distinct from visceral and yolk sac endoderm (VE/YE). Exmes upregulate COL6A1 while downregulating FOXA1, whereas VE/YE exhibit the opposite pattern[71]. These findings support the view that Exmes are hypoblast-related but molecularly divergent lineage, rather than a simple extension of VE/YE identity.
In vitro evidence
Stem cell-based models have provided important mechanistic insights into the specification and maintenance of Exmes. Directed differentiation studies demonstrate that modulation of BMP, WNT and Nodal signalling is sufficient to induce Exmes-like populations from human pluripotent stem cells (hPSCs). For example, primed hESCs can be differentiated into early Exmes-like cells through coordinated activation of BMP and WNT pathways alongside modulation of Nodal signalling[10]. Similarly, naïve human pluripotent stem cells can be converted into self-renewing Exmes populations through inhibition of Nodal signalling and GSK3β, with sustained proliferation requiring activation of mTOR and BMP pathways. Consistent with this, inhibition of BMP signalling reduces SMAD1/5/9 phosphorylation and markedly impairs Exmes expansion[74].
Additional studies further indicate that activation of WNT signalling can generate Exmes populations with hematopoietic potential, characterized by expression of MEF2C, CD34, PECAM1 and CDH5[75]. Notably, multiple induction systems applied to naïve hESCs (AIC-N), naïve pluripotent stem cells and hiPS cells all yielded Exmes-like cells with transcriptional signatures closely resembling hypoblast-derived Exmes
Summary of markers for the lineages in human embryos (Trophoblast, Epiblast, Hypoblast), for Exmes in embryo models supporting different Exmes origin hypotheses, and for EXMC following gastrulation
| Cell | Markers | Period | From |
| Trophoblast | GATA3, KRT7[72,73,77], GATA2[73], CDX2[71,76] | ||
| Epiblast | POU5F1 (OCT4)[73], NANOG, SOX2, PRDM14[71] | ||
| Hypoblast | GATA4, GATA6, PDGFRA, SOX17[7,72], DPPA3[73,76], APOA1[7,62,73], CER1, CXCR4, POSTN, FGFR1, NODAL, OTX2[7], FOXA2[62,76], IHH, FOXA1, HNF4A[62] YS.endoderm markers: TTR[7,73], BMP6, RBP4, GC, APOB, PDGFRA[7] AVE markers: LHX1, HHEX, EOMES, GSC, CER1, LEFTY1, LEFTY2, NOG[62] | ||
| Primitive streak | TBXT, MIXL1, EOMES, HAND1 [7,76] | ||
| Pre-gastrulation | |||
| Hypothesis of epiblast -derived | |||
| Exmes | Early: LUM, NID2, FOXF1, VIM, POSTN, GATA4, ANXA1, COL4A1, COL4A2, BMP4 Late: LUM, NID2, VIM, POSTN, PTX3, NR2F2 | 8, 13, 18 days (early) and 70 days (late) induced by ASECRiAV medium | Naïve hPSCs[74] |
| NID2, FOXF1, VIM, ANXA1 (FOXA2 and LUM negative) | 8, 24 days induced by RACL medium | ||
| MIXL1, MESP1, ETS1, PODXL, EPHA4, ZIC3 | 10 days | Cynomolgus macaque embryos (in vitro)[69] | |
| PODXL, ETS1, PITX1, LUM, NID2 | 12 days | ||
| PITX1, LUM, NID2, FOXF1, PDGFRA, HAND1 | 14 days | ||
| Hypothesis of hypoblast -derived | |||
| Exmes | GATA4, GATA6, COL6A1 | E13-E17 (early post-implantation) | Cynomolgus monkeys (in vivo and in vitro)[71] |
| GATA4, GATA6, PDGFRA, NID2, BST2 | 6 days induced by RCL medium | Naïve PS (in vitro)[72] | |
| GATA4, GATA6, CDH2, KDR, VIM, FLT1, COL3A1, SNAIL | 4 days | Naïve hESCs (AIC-N)[10] | |
| EMT-related genes (ISM1, SNAI2, SPOCK1); ECM genes (ADAM12, COL5A1, POSTN) | 0-2 days | PrE-like cell line (naïve extraembryonic endoderm) (in vitro)[61] | |
| GATA6, PLAGL1, PLVAP, GATA2, KLF1 BST2, COL1A1, COL1A2, COL2A1 | 5 days | hiPS (in vitro)[78] | |
| GATA4, GATA6, PDGFRA, FN1, COL4A1, COL6A1, VIM, SNAI2, KDR, FLT1, MEIS2, COL3A1, NID2, FOXF1 | E10-14 | Human (in vitro)[62] | |
| HAND1, VIM, SNAI2, KDR, FLT1, MEIS2, COL3A1, COL6A1, FOXF1, POSTN | 8 days | E-assembloid (AIC-N hESC & xEMs)[62] | |
| VIM, KDR, NID2, COL1A1, COL3A1, LUM, BST2, POSTN, DCN | 7 days | hpESCs (in vitro)[75] | |
| Post-gastrulation | |||
| EXMC | GATA6, BMP4, HAND1, SNAI2, FOXF1, POSTN, ANXA1 | CS7 | Human (in vivo)[5] |
| BMP4, LUM, MYL7 | CS7 | Human (in vivo)[79] | |
| LUM, POSTN, IGF2 | CS8 | Human (in vivo)[80] | |
| DEPTOR, FBN2, FRMD4B, HAND2, OAF | CS7-CS8c | Monkey blastocyst (in vitro)[81] | |
| GATA4, GATA6, COL1A1, COL1A2, COL3A1, TGFBI, LUM, TIMP1, DCN, IGFBP3, TAGLN | 9-20 days | Cynomolgus monkeys (in vitro)[73] | |
| HAND1, SNAI2, HGF, PDGFRA, WNT5A, FOXF1, BMP4, VIM, IL7R, GNG11 | CS5, 6, 7 | Marmoset (in vivo)[7] | |
| POSTN, COL6A1 | 17 days | Cynomolgus monkey (in vitro)[76] | |
| Connecting stalk markers: CDX1, CDX2 | hPSCs (in vitro)[82] | ||
| CDX1, CDX2 | CS7 | Human (in vivo)[79] | |
| CDX2, CDH2, LCN15 | CS8 | Human (in vivo)[80] | |
Among these models, the model proposed by Pham is particularly informative regarding dual contributions from epiblast- and hypoblast-related routes to extraembryonic mesoderm. In this system, naïve hPSCs are first directed through an epiblast-like state (expressing POU5F1/OCT4 and NANOG) and then induced, using ASECRiAV conditions, to differentiate into extraembryonic mesoderm cells expressing POSTN, VIM, NID2, LUM, FOXF1, ANXA1 and DCN. In parallel, conversion of naïve hPSCs to PrE using RACL generates extraembryonic mesoderm cells expressing NID2, FOXF1, VIM and ANXA1. Notably, this hypoblast-biased route also appears to transit through an intermediate epiblast-like phase, as indicated by transient expression of POU5F1, NANOG, SOX2, DPPA2, GDF3, ZNF208 and ZNF729[74]. Together, these data suggest the notion that Exmes can arise through multiple developmental trajectories, potentially reflecting the temporally distinct contributions inferred in vivo.
Molecular and functional features of Exmes
Beyond lineage origin, Exmes exhibit a distinct molecular identity consistent with their structural and regulatory roles in early development.
Exmes form an extracellular matrix (ECM)-rich mesenchymal stroma, characterized by high expression of ECM components[61,71,73,75]. They produce abundant fibronectin (FN1), collagens (e.g. COL1A1, COL6A1) and laminin-111, which accumulate prominently around the embryonic-extraembryonic interface, particularly in the connecting stalk region[71]. In trilaminar cynomolgus monkey embryos derived from naïve ESCs, Exmes show upregulation of EMT-related ECM genes such as POSTN and COL6A1[76].
Moreover, Exmes display a mesenchymal phenotype marked by expression of PDGFRA and activation of EMT-related transcriptional programme[7,61,71,73]. For instance, in early post-implantation cynomolgus embryos (CS5), Exmes show high expression of SNAI2 and HGF[7], consistent with activated EMT and invasive behaviour during early development.
In addition to their structural role, Exmes function as a signalling hub that interfaces between embryonic and extraembryonic tissues via ligand-receptor networks. They express key components of TGF-β signalling which regulates EMT, including TGFB1, ZEB2, SMAD3, SNAI2 and VIM[73], as well as ligands associated with WNT and BMP pathways[7]. This suggests that Exmes act as a signalling hub to modulate epiblast, hypoblast and trophoblast compartments.
Taken together, these features support a model in which early Exmes are primarily generated from hypoblast-related cells via EMT and robust ECM production. As development proceeds and gastrulation begins, the bilaminar disc transitions into a trilaminar structure. The newly formed mesoderm layer expands extraembryonically, and EXMC derived from the primitive streak infiltrate regions previously occupied by Exmes. At this stage, a mixed Exmes/EXMC population builds the stromal, vascular and immunological architecture of extraembryonic tissues. It is this composite Exmes/EXMC compartment that ultimately underpins the diverse structural and functional roles traditionally attributed to the extraembryonic mesoderm.
EXMC AFTER GASTRULATION
Human gastrulation takes place between days 14 and 21 post-fertilization (CS7-CS9)[5]. During this interval, the bilaminar disc is transformed into a trilaminar embryo as the epiblast gives rise to the three germ layers: ectoderm, mesoderm and endoderm[5].
In the mouse, it is well established that, after gastrulation begins at approximately E6.5, EXMC arise from mesodermal cells that proliferate and migrate outwards from the primitive streak[13]. By contrast, in primate embryos, the corresponding extraembryonic region is already occupied by a fully formed Exmes population during the early post-implantation, pre-gastrulation period. Thus, primate Exmes are generally considered to have a distinct origin from rodent EXMC. Nevertheless, once gastrulation is underway and the trilaminar embryo has formed, PS-derived mesoderm is likewise expected to proliferate and extend into the extraembryonic space, in a manner analogous to the mouse, thereby generating EXMC that colonize territories previously filled by Exmes.
EXMC in extraembryonic tissues after the onset of gastrulation
A current working model therefore posits that, in early primate embryos, the extraembryonic compartment is initially filled by Exmes. Following gastrulation, at the trilaminar germ-disc stage, newly generated mesoderm gives rise to EXMC that migrate into and intermix with Exmes in the extraembryonic region, producing a composite Exmes/EXMC population. Classical anatomical work in lemur embryos by Hill indicated that primate EXMC extend from the primitive streak as an outward continuation of the gastrulation-derived mesoderm[63]. More recently, studies of human CS8 embryos have shown that, under graded signalling along the A-P axis and the primitive streak, distinct mesodermal subpopulations occupy specific positions along the streak. Within this pattern, EXMC progenitors are localized to the outermost domain of the embryonic disc, at the interface with the extraembryonic region[79].
Within this framework, EXMC can be spatially subdivided into at least two functionally distinct subpopulations, whose differentiation trajectories depend on their microenvironment [Figure 7]. EXMC that spread ventrally into the yolk sac domain form the splanchnic EXMC[67] surrounding the yolk sac; this population contributes to the formation of yolk sac blood islands and drives the earliest wave of extraembryonic hematopoiesis. By contrast, EXMC that expand toward the outer surface of the amnion and into the trophoblast villous core give rise to the stromal compartments of the amnion and placental villi, respectively[67].
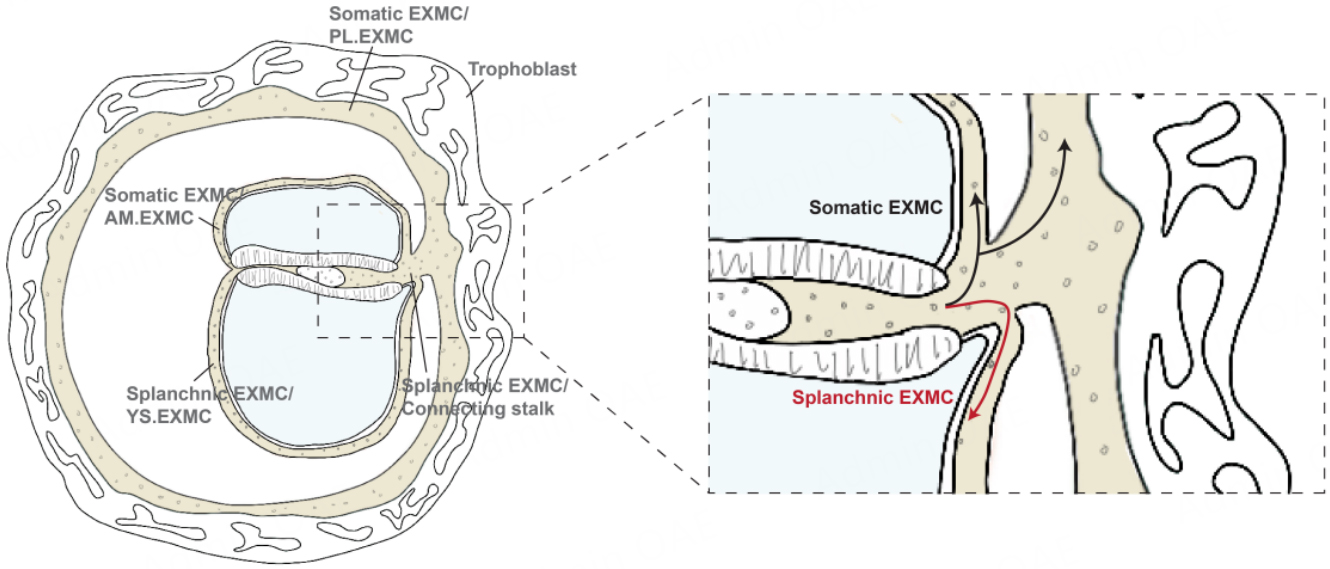
Figure 7. Spatial heterogeneity exists within the EXMC, comprising splanchnic EXMC in the extraembryonic space of the yolk sac and somatic EXMC in the extraembryonic space between the amniotic cavity and trophoblast. Different EXMC cell populations also exhibit functional differences. Image created by Adobe Illustrator.
Signalling mechanisms regulating EXMC specification and expansion
In primates, similar to rodents, BMP4 plays a central role in the induction and differentiation of EXMC. BMP4 targets SLUG and MSX2 to amplify EMT programmes and promote the specification of mesodermal lineages, including EXMC[83]. BMP4 also supports the renewal and maintenance of EXMC through SMAD1/5/9 phosphorylation[74]. Synergistic interactions between BMP4 and other signalling pathways have been demonstrated in in vitro systems. For example, BMP and FGF2 cooperate to drive hESC differentiation towards mesoderm and EXMC via BRA and CDX2[84]. Sustained BMP4 and WNT signalling can inhibit endoderm differentiation and instead bias hESCs towards EXMC-like fates[82,85].
BMP signalling is also essential for amnion development. Amniotic epithelial cells express multiple BMP ligands, and BMP4 acts downstream of ISL1; loss of ISL1 in AmE perturbs amnion-derived signalling, reduces BMP4 expression and leads to abnormal mesoderm differentiation[69].
Recent single-cell analysis of human early-pregnancy (5-9 weeks) amnion by Hu and colleagues resolved transcriptionally distinct amniotic epithelial cells (GABRP, VTCN1) and amniotic mesenchymal cells (MGP, VIM, POSTN). EMT-associated transcription factors (SNAI2, TWIST1, TWIST2) were enriched in the mesenchymal compartment, and high SNAI1 expression was observed in amniotic epithelial stem cells poised near the mesenchymal trajectory. These findings suggest that amniotic mesenchymal cells can arise from amniotic epithelium via EMT, with epithelial stem cells functioning as a transitional population[86]. By contrast, in earlier CS8 human embryos, amniotic EXMC have been proposed to derive from EXMC progenitors generated during gastrulation that subsequently migrate towards the amnion[79]. Whether amniotic EXMC are unified in origin or reflect multiple temporally and spatially distinct progenitor pools therefore remains an open question.
EXMC are closely related to hematopoiesis
The extraembryonic mesodermal core of the placental villi begins to develop capillary-like structures[34], reminiscent of yolk sac blood islands, around 18-20 days. By 18 days, primitive myeloid and erythroid cells (HBCs and nucleated erythrocytes) can already be detected within the placenta, and by the fourth week the villous architecture is well established, containing fetal vessels and abundant HBCs[87]. This hematopoietic activity, which closely mirrors that observed in yolk sac blood islands, is consistent with a mixed state in which splanchnic and somatic EXMC populations interdigitate within the villous core. In this scenario, the hematopoietic contribution of the placental villi would derive from splanchnic EXMC intermixed with somatic EXMC, providing a mechanistic explanation for how villi are able to generate blood vessels and blood cells before the umbilical cord establishes a definitive vascular connection between the embryo and the placenta. Intriguingly, amniotic epithelial cells also exhibit the capacity to differentiate into macrophages. This transition is accompanied by downregulation of epithelial markers (EPCAM, GABRP, IGFBP3, CLDN6) and upregulation of macrophage markers (CD14, CD163, CD36, MRC1)[86]. Together with their EMT potential, these data raise the possibility that the amnion, like the yolk sac, may serve as an in situ extraembryonic hematopoietic niche.
CONTRIBUTION OF EXMES/EXMC TO EARLY EMBRYONIC HEMATOPOIESIS
Early embryonic hematopoiesis is one of the most striking functional contributions of Exmes/EXMC. By 5-6 weeks of gestation, substantial numbers of CD34+ cells are present in the placenta[88]; yet whether the placenta can sustain in situ hematopoiesis remains debated. There are three canonical hematopoietic waves: primitive wave (CS7-8), transient-definitive wave (CS8-9), and aorta-gonad-mesonephros (AGM)-derived definitive wave (CS13-15)[89]. Evidence from early primate embryos suggests that hematopoietic activity may begin even before these stages. Moreover, studies of HBCs in placental villi prior to CS10[90,91], before the onset of cardiac circulation, are consistent with a possible early, in situ hematopoietic contribution from extraembryonic tissues, although the developmental independence of these cells from yolk sac hematopoiesis remains to be clarified.
Angiogenesis of the connecting stalk and its hematopoietic potential
Hematopoietic potential of the allantois in rodents
In rodents, the allantois is not merely an excretory and respiratory appendage but also a source of hematopoietic and endothelial progenitors[92]. Pre-fusion allantoic explants can generate erythroid and myeloid cells in vitro[93], demonstrating an intrinsic hematopoietic potential before vascular connection with the yolk sac. Allantoic hematopoiesis is Runx1-dependent: Runx1-deficient allantois fails to produce hematopoietic cells[94]. Vascularization within the allantois exhibits a distal-proximal polarity, with a branched vascular plexus distally and straight umbilical arteries proximally, ensuring appropriate docking with the chorion and correct vascular patterning[48].
Formation and patterning of the connecting stalk in primates
In primate embryos, rapid expansion of Exmes drives the emergence of the connecting stalk. Between days 13 and 16, Exmes initially cover the outer surface of the amnion and yolk sac[3]. Because Exmes cannot keep pace with this expansion, the embryo grows predominantly towards the anterior side, and Exmes condenses into a stalk-like structure that remains tethered at the posterior pole[35]. The connecting stalk later differentiates into extraembryonic structures including the umbilical cord and allantois, which link to the placenta and mediate maternal-fetal exchange. Functionally, the connecting stalk establishes a conduit between embryonic and extraembryonic tissues that enables the circulation of fetal blood between intraembryonic and extraembryonic compartments. In addition, the connecting stalk has been proposed to act as a signalling centre that helps pattern the embryonic A-P axis, potentially through cell migrations that occur after gastrulation. In this context, it is plausible that, following gastrulation, mesenchymal cells in the stalk region become admixed with EXMC generated from mesoderm, mirroring the mixed Exmes/EXMC state inferred in other extraembryonic territories.
Single-cell transcriptomic analyses of human CS7-CS8 embryos identified a population of cells at the posterior mesoderm that expressed markers associated with the connecting stalk, including CDX1, CDX2, CDH2 and LCN15[79,80]. CDX2 also serves as a molecular marker for the posterior EXMC, which includes the connecting stalk. Knockout of CDX1/CDX2 leads to proliferation defects in EXMC and delayed allantoic development[82,95]. Under normal conditions, FGF and BMP signalling at the posterior pole suppress Nodal/ACTIVIN activity, downregulate NANOG and upregulate BRA and CDX2, thereby promoting posterior fates while inhibiting anterior lateral plate mesoderm, definitive endoderm programmes, and cardiac development. In the anterior domain, NANOG antagonizes CDX2 expression[82].
Gene perturbation studies of CDX2 and TBXT further indicate that EXMC downregulate the WNT inhibitor DCDC2, and that angiogenesis is attenuated through modulation of VEGF signalling-via downregulation of ANK3 and ANGPT1 and upregulation of TMSB10[95]. These data link posterior EXMC identity to vascular patterning and hematopoietic competence in the connecting-stalk and allantoic region.
Villous filling and angiogenesis within Exmes
Hematopoietic potential of chorionic villi in rodents
In the mouse, the placenta is not only a niche for hematopoietic stem cells (HSCs)[96,97] but also an active hematopoietic organ[97-99]. Early studies identified functional hematopoietic cells with immune capabilities in the placenta[98]. Immature hematopoietic cells expressing CD41, Runx1, c-Kit, CD31, CD34 and Sca-1 have been detected in the labyrinthine zone[100]. The number of HSCs in the placenta peaks at E12.5-E13.5, reaching about 15-fold the number in the AGM region[96]. Importantly, hematopoietic cells capable of autonomous generation are present in the chorion before chorio-allantoic fusion[93,94], indicating that chorionic hematopoiesis does not merely reflect colonizing from the yolk sac or circulation. The spatiotemporal expression of Runx1 in the chorion and at the chorion-allantois interface resembles that of integrin α4[94], a key mediator of chorio-allantoic fusion[42,51], suggesting that Runx1 may coordinate both fusion and the establishment of placental hematopoietic niches.
Villous expansion and angiogenesis in primate Exmes
In primates, the expansion of Exmes play an inductive role in chorionic villus development. Villous morphogenesis begins at approximately days 11-13, when the chorion differentiates from trophoblast. pSTB invades the decidua, while pCTB proliferates to form primary villi, consisting of pCTB cores surrounded by multinucleated pSTB [Figure 3A and B]. Around days 13-15, Exmes infiltrate these primary villi to generate the first mesenchymal cores, filled with a reticular matrix and constituting secondary villi[34]
Paracrine factors secreted by trophoblast cells are widely thought to induce villous mesenchymal differentiation and initiate vascular development[101,102]. The first fetal capillaries appear around days 18-20. By approximately day 22, angiogenic cells and endothelial cords interconnect through loose junctions and elongate along the villous axis, ultimately giving rise to branched vessels[101]. Perivascular and smooth muscle cells then encase these primitive tubes to form contractile vessels, which mature by approximately 28 days and contribute to tertiary villi. Chorionic villi near the connecting stalk further differentiate into the definitive placenta, which serves as the primary maternal-fetal interface.
Within this framework, Exmes expansion and differentiation provide both the scaffold and the cellular substrate for villous angiogenesis and early placental hematopoiesis, helping to explain how villi can generate blood vessels and blood cells before the umbilical circulation is fully established.
Hypothesis of early hematopoiesis and placental in situ hematopoiesis in primate embryos
Exmes are associated with embryonic hematopoiesis
In mammals, the first transient blood cells are generated in the yolk sac[103]. Early yolk sac hematopoiesis is classically divided into two waves. A primitive wave, initiating before or around CS7, produces early erythromyeloid and megakaryocytic progenitors, whereas a subsequent definitive wave at CS8-CS9 generates erythromyeloid and lymphoid-lineage cells. Blood islands on the yolk sac function as the first recognizable hematopoietic centres[4] [Figure 8]. These blood islands arise from solid clusters of mesenchymal cells aggregating near the endoderm within the yolk sac mesoderm-regarded as the anlage of blood islands-and are subsequently induced to differentiate by adjacent endodermal cells [Figure 8]. Structurally, blood islands consist of flattened endothelial progenitors encasing primitive hematopoietic progenitors, forming a characteristic hemato-endothelial unit[4] [Figure 8]. The flattened endothelial progenitors of blood islands differentiate into hemogenic endothelium through hemogenic specification, which subsequently gives rise to hematopoietic stem and progenitor cells via the endothelial-to-hematopoietic transition pathway[104]. Rapid expansion and remodelling of these units give rise to an extensive vascular plexus that envelops the yolk sac in a complex arborizing network. The primitive wave of yolk sac hematopoiesis satisfies the early hematopoietic demands of the embryo, and with the onset of cardiac contractions at CS10[91], yolk sac-derived blood cells are disseminated throughout the embryonic body.
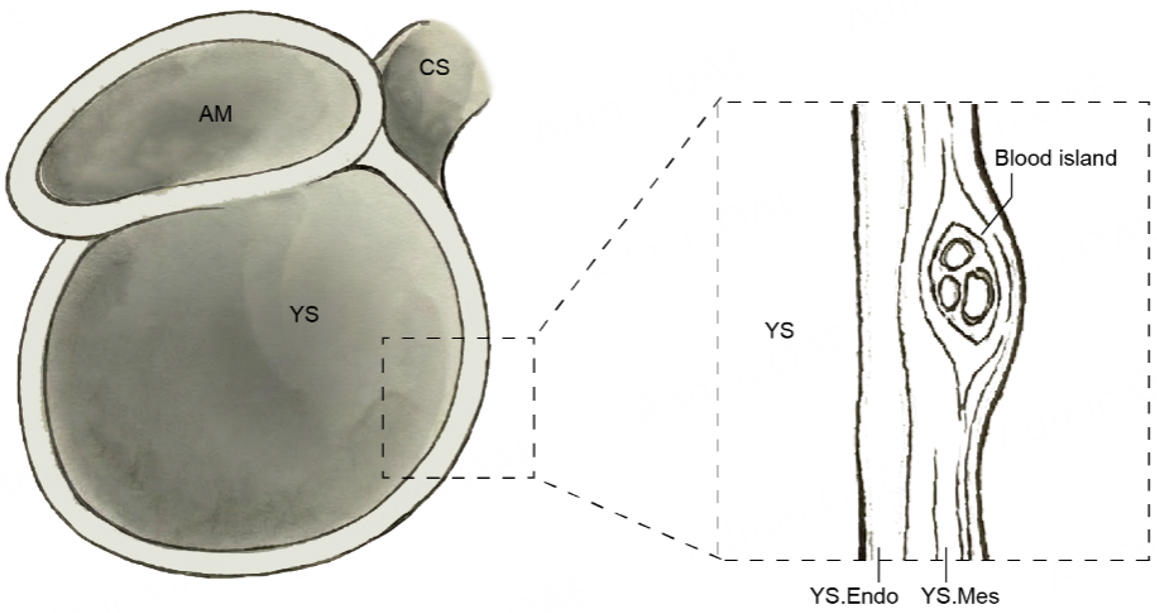
Figure 8. Blood islands begin as solid mesenchymal cell clusters aggregated within the yolk sac mesoderm. Structurally, they consist of flattened endothelial progenitor cells surrounding primitive hematopoietic progenitor cells. AM: Amnion; YS: yolk sac; CS: connecting stalk; YS.Endo: yolk sac endoderm; YS.Mes: yolk sac mesoderm. Image created by Adobe Illustrator.
In vitro models, Exmes induced from PrE recapitulate key features of this early extraembryonic hematopoietic niche. In addition to upregulating EMT- and ECM-associated genes, these Exmes populations show increased expression of angiogenesis- and hematopoiesis-related genes such as KDR and DPP4, as well as the placental angiogenesis-associated gene EMP2, from around day 8 of induction[61]. In another human embryo-like model, the heX-embryoid-which contains yolk sac hematopoietic tissue and an extraembryonic niche-a distinct Exmes population has been identified. These cells express high levels of the surface marker BST2 and ECM components, while co-expressing endothelial genes (CD34, PECAM1) and hematopoietic genes (GATA1, KLF1, ZEB1, CD235)[78]. In addition, a population of endothelial cells expressing CD34, CDH5, ERG and GATA2, but lacking CD73, has been observed, consistent with an EHT toward progenitor cells[78]. Together, these findings demonstrate that Exmes derived from yolk sac mesoderm make a substantial contribution to the primitive wave of hematopoiesis during early development and highlight the capacity of in vitro models to capture early extraembryonic hematopoietic programmes.
Hypothesis of placental in situ hematopoiesis in primate embryos
The placenta is a pivotal extraembryonic organ required for gas exchange, nutrient supply and endocrine support of the developing embryo. Its function is critically dependent on the timely establishment of maternal-fetal circulation. The basic villous architecture of the human placenta is in place by approximately 4 weeks of gestation[15], coinciding with the period during which Exmes fill the villous cores and undergo differentiation and maturation. However, the temporal uncoupling between the appearance of HBCs in human placental villi at approximately CS8[90], well before the onset of embryonic heart beat and the establishment of a functional fetoplacental circulation at CS10[91], has intensified interest in whether Exmes within the villous stroma support an early, in situ hematopoietic programme independent of fetal blood cellular colonization.
Hematopoietic activity in the placenta
The placenta is well established as a niche for the maturation of hematopoietic progenitors: throughout gestation, it harbours both hematopoietic progenitor cells and HSCs[96,105,106]. Macrophages arise earlier than HSCs and derive from the first wave of yolk sac primitive hematopoiesis, subsequently colonising tissues as tissue-resident macrophages (TRMs)[107]. In the placenta, HBCs function as TRMs and are essential for maintaining normal placental development and pregnancy homeostasis[107]. A recent study reported that two populations of Exmes/EXMC contribute respectively to placental mesenchymal cells and to HBCs, erythrocytes, and endothelial cells[108].
HBCs express CD14 and CD68, together with receptors including CD163, CD206, CD64 and FOLR2[109]. In early pregnancy, HBCs derived from primitive hematopoiesis lack expression of the classical macrophage marker HLA-DR[109], a phenotype linked to epigenetic silencing of CIITA (class II transactivator)[110].
Studies have shown that fetal-derived CD34+, CD43+ and FXIII/CD68+ macrophage precursors are already present in villous cores before the establishment of placental circulation[111]. Single-cell and functional assays indicate that early human HBCs share transcriptional signatures with yolk sac macrophages and express proliferation-associated genes such as Ki67. These findings suggest that early placental macrophages originate from primitive hematopoiesis and are capable of local proliferation and self-renewal within the placenta[109].
HBCs heterogeneity
HBCs can be classified into two major subpopulations based on their origin in early and late pregnancy. HBCs in early pregnancy, which arise either from villous mesenchymal stem cells or from monocyte precursors originating in the yolk sac, are potentially associated with tissue development and morphogenesis; HBCs in late pregnancy, which derive from definitive HSCs are likely involved in hematopoiesis and immune functions[112].
These subtypes exhibit distinct expression patterns: CD68 is expressed throughout gestation; CD1, CD4, CD11c, CD14, CD16, CR3 and CXCR4 are enriched in early and late pregnancy; ACP and G6PD are highly expressed in early to mid- pregnancy; CD163, CD209, and HLA-DQ are specifically highly expressed in late pregnancy[113].
The origin of HBCs in primates
The precise origin of HBCs in early pregnancy, whether primarily placental or yolk sac-derived, remains unresolved. Spatiotemporally, the very early appearance of HBCs is congruent with their putative functions: they must differentiate and mature rapidly to support tissue remodelling, repair and angiogenesis, rather than waiting for yolk sac hematopoiesis and cardiac-driven circulation to be fully established.
At day 18, HBCs have been observed within placental villi[114,115]. Importantly, the appearance of HBCs precedes the onset of fetal circulation at CS10[91], when blood flow to the placenta is not yet established, making large-scale migration of macrophage precursors from the yolk sac into villi mechanistically challenging.
It is also debated whether early HBCs arise from a yolk sac-independent primitive programme within the placenta or from yolk sac-derived primitive progenitors that colonize the villi. In the mouse, HBC-like placental macrophages can arise from erythro-myeloid progenitors (EMPs) of primitive hematopoiesis. Analogously, a population of CD34+ CD43+ HLF+ placental erythro-myeloid progenitors (PEMPs) has been identified in early human placenta, transcriptionally reminiscent of murine EMPs. PEMPs express high levels of CD45 and c-KIT and, in culture, give rise to HLA-DR⁻ HBCs, allowing reconstruction of their differentiation trajectory[110].
Current hypotheses regarding the origin of HBCs in primate placenta therefore include: (i) differentiation from villous mesenchymal cells; (ii) migration and differentiation of yolk sac derived mononuclear precursors; and (iii) recruitment of yolk sac-derived macrophages after the onset of embryonic circulation. HBCs of distinct developmental origin are likely to perform non-overlapping functions, contributing to HBCs heterogeneity.
Disentangling the relative contributions of placental and yolk sac-derived hematopoiesis remains challenging, not only because of technical limitations in tracing early macrophage ontogeny, but also because very early-stage human specimens are rare and human and murine placentas differ substantially in structure, architecture and timing.
DISCUSSION
Exmes/EXMC have received limited attention in mainstream embryology, which has traditionally focused on patterning of the embryonic germ layers. Comparative studies in rodents, non-human primates and humans[3,4,13], together with stem cell-based embryo models, now position Exmes/EXMC within a revised framework. Rather than passive supportive stroma, Exmes/EXMC constitute a dynamic and heterogeneous continuum that scaffolds the amnion, yolk sac, connecting stalk and placenta. Concurrently, these cells support early hematopoiesis, vascular patterning and maternal-fetal communication during the brief but decisive peri-implantation to early-organogenesis window[1].
Cross-species studies indicate that primate extraembryonic mesoderm is neither temporally nor molecularly uniform. In rodents, classical fate mapping supports a relatively simple model in which extraembryonic mesoderm arises from posterior epiblast cells that ingress through the primitive streak during gastrulation and subsequently expand into extraembryonic territories[2]. By contrast, primate embryos already harbour mesenchymal-like Exmes within the chorionic cavity and around the primary yolk sac before overt mesoderm formation[3,4]. Gastrulation-derived EXMC arrive later into the same domain. Histological, single-cell and stem cell-based data support a two-step model: an early, pre-gastrulation Exmes population transcriptionally closer to hypoblast/PrE, followed by a wave of EXMC produced by mesoderm during gastrulation[67]. Explicitly distinguishing pre-gastrulation Exmes from gastrulation-derived EXMC helps reconcile long-standing discrepancies between classical descriptions and modern molecular data, while emphasizing that these populations later intermingle.
The developmental origin of early Exmes in primates nevertheless remains unresolved. Early morphological work proposed a trophoblastic origin for the reticular mesenchyme lining chorionic and villous cavities[41]. However, the presence of a basal lamina separating trophoblast from villous mesenchyme[65] and the expression of ICM-associated transcription factors in Exmes-like cells argue against this view[67]. Recent in vivo and in vitro datasets instead implicate the ICM. Two broad scenarios have emerged. Hypoblast-focused models[71-73] posit that yolk sac endoderm delaminates and undergoes epithelial-mesenchymal transition to yield Exmes. Epiblast-focused models propose that epiblast-derived cells contribute extraembryonic mesoderm earlier than previously appreciated[3,68,69]. At present, both models rest on marker co-expression and trajectory analysis rather than definitive lineage tracing. Disentangling hypoblast-, epiblast- and mesoderm-derived contributions will require genetically based fate mapping and temporally controlled perturbations in non-human primates. High-fidelity human stem cell-based embryo models that robustly generate extraembryonic lineages will also be necessary.
Functionally, Exmes/EXMC serve as organizers of extraembryonic vascular and hematopoietic programmes. In primate yolk sacs, Exmes/EXMC aggregates generate blood-island-like structures, endothelial networks and early erythro-myeloid progenitors before the establishment of robust cardiac contraction and systemic circulation[4]. In nascent placental villi, Exmes differentiate into vascular endothelium, perivascular stroma and HBCs, the TRMs of the placenta. HBCs appear in human villi by approximately embryonic day 18 (CS8)[114,115], whereas a functional fetoplacental circulation driven by embryonic cardiac contractions is not established until around CS10[91]. This temporal uncoupling raises the question of whether villous Exmes support an early, in situ haematopoietic programme that is at least partially independent of yolk sac-derived seeding. The identification of placental erythro-myeloid progenitors and proliferative HBCs with primitive signatures is consistent with such a model. However, whether these cells arise from a placenta-intrinsic programme or from very early yolk sac progenitors remains unclear.
Clinically, Exmes/EXMC biology is poised to illuminate the earliest phases of human pregnancy, during which many miscarriages, placental disorders and long-term health trajectories are established. Subtle defects in Exmes deployment, matrix organization, vascularization or macrophage programmes may compromise the competence of extraembryonic tissues long before symptoms become apparent. Linking human genetic variation and early pregnancy outcomes to disruption of Exmes/EXMC programmes may identify targets for intervention in early pregnancy. As ethical and regulatory frameworks for human embryo research and embryo-like models evolve, bringing Exmes/EXMC into the conceptual centre of early mammalian development will be essential for a truly integrated view. Such integration will ensure that the construction of extraembryonic tissues is recognized as equally fundamental to successful gestation as the patterning of the embryonic germ layers.
CONCLUSION
In this review, we review current evidence of Exmes/EXMC during early mammalian development, emphasizing their conserved functional importance and divergent developmental origins across species. Evidence from classical embryology, genetic studies, and recent single-cell and stem cell-based models consistently establishes Exmes/EXMC as essential contributors to the formation and organization of extraembryonic tissues, including the amnion, yolk sac, connecting stalk, and placenta. Through their roles in extracellular matrix deposition, epithelial support, and paracrine signaling (including BMP, WNT, and TGF-β pathways), these cells establish the biomechanical and molecular framework necessary for axis formation, and tissue morphogenesis. Moreover, they act as critical regulators of early vascularization and hematopoiesis, contributing to yolk sac blood island formation, placental villous development, and potentially to in situ hematopoietic niches prior to the establishment of embryonic circulation.
A key conclusion is the distinction between rodent and primate developmental strategies. In rodents, EXMC arise in a well-defined manner from gastrulation-stage mesoderm. In contrast, primate embryos exhibit earlier Exmes prior to gastrulation, followed by the addition of mesoderm-derived EXMC. This developmental asynchrony underscores a lineage continuum in primates that is temporally expanded and structurally more complex.
Importantly, advances in stem cell-based embryo models and multi-omics approaches have begun to bridge gaps imposed by limited access to early primate embryos, enabling more precise characterization of Exmes/EXMC. These systems are now necessary for reconstructing early lineage and functional properties. Highlighting Exmes/EXMC as a central, yet previously underappreciated, lineage in early development, provides a conceptual framework for future studies aimed at understanding extraembryonic tissue biology and its relevance to human reproductive health.
DECLARATIONS
Authors’ contributions
Conceptualized the idea, and oversaw the overall project: Xiao Z
Performed manuscript writing, image drawing, and editing with input and feedback from all authors:
Availability of data and materials
Not applicable.
AI and AI-assisted tools statement
Not applicable.
Financial support and sponsorship
This work was supported by the National Key Research and Development Program of China (2024YFA1107500 and 2024YFA1107502).
Conflicts of interest
Both authors declared that there are no conflicts of interest.
Ethical approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Copyright
© The Author(s) 2026.
REFERENCES
1. Nehme E, Panda A, Migeotte I, Pasque V. Extra-embryonic mesoderm during development and in in vitro models. Development. 2025;152:DEV204624.
2. Thowfeequ S, Hanna CW, Srinivas S. Origin, fate and function of extraembryonic tissues during mammalian development. Nat Rev Mol Cell Biol. 2024;26:255-75.
3. Luckett WP. Origin and differentiation of the yolk sac and extraembryonic mesoderm in presomite human and rhesus monkey embryos. Am J Anat. 2005;152:59-97.
4. Ross C, Boroviak TE. Origin and function of the yolk sac in primate embryogenesis. Nat Commun. 2020;11:3760.
5. Tyser RCV, Mahammadov E, Nakanoh S, Vallier L, Scialdone A, Srinivas S. Single-cell transcriptomic characterization of a gastrulating human embryo. Nature. 2021;600:285-9.
6. Xiang L, Yin Y, Zheng Y, et al. A developmental landscape of 3D-cultured human pre-gastrulation embryos. Nature. 2019;577:537-42.
7. Bergmann S, Penfold CA, Slatery E, et al. Spatial profiling of early primate gastrulation in utero. Nature. 2022;609:136-43.
8. Srivatsan SR, Regier MC, Barkan E, et al. Embryo-scale, single-cell spatial transcriptomics. Science. 2021;373:111-7.
9. Ren H, Jia X, Yu L. The building blocks of embryo models: embryonic and extraembryonic stem cells. Cell Discov. 2025;11:40.
10. Niu B, Wang D, Hu Y, et al. Deciphering signaling mechanisms and developmental dynamics in extraembryonic mesoderm specification from hESCs. Nat Commun. 2025;16:4688.
11. Okubo T, Takashima Y. Exploring the human extraembryonic mesoderm using naive pluripotent stem cells. Cell Stem Cell. 2022;29:1290-1.
12. Karvas RM, Zemke JE, Ali SS, et al. 3D-cultured blastoids model human embryogenesis from pre-implantation to early gastrulation stages. Cell Stem Cell. 2023;30:1148-65.e7.
13. Shahbazi MN, Zernicka-goetz M. Deconstructing and reconstructing the mouse and human early embryo. Nat Cell Biol. 2018;20:878-87.
14. Siriwardena D, Boroviak TE. Evolutionary divergence of embryo implantation in primates. Phil Trans R Soc B. 2022;377:20210256.
15. Hemberger M, Hanna CW, Dean W. Mechanisms of early placental development in mouse and humans. Nat Rev Genet. 2019;21:27-43.
16. Bedzhov I, Zernicka-Goetz M. Self-organizing properties of mouse pluripotent cells initiate morphogenesis upon implantation. Cell. 2014;156:1032-44.
17. Kim YS, Fan R, Kremer L, et al. Deciphering epiblast lumenogenesis reveals proamniotic cavity control of embryo growth and patterning. Sci Adv. 2021;7:eabe1640.
18. Yamamoto M, Saijoh Y, Perea-Gomez A, et al. Nodal antagonists regulate formation of the anteroposterior axis of the mouse embryo. Nature. 2004;428:387-92.
19. Chen D, Claussen NH, Titus S, et al. Basement membrane perforations guide anterior-posterior axis formation. Nat Commun. 2025;16:6763.
20. Morris SA, Grewal S, Barrios F, et al. Dynamics of anterior-posterior axis formation in the developing mouse embryo. Nat Commun. 2012;3:673.
21. Chuva De Sousa Lopes SM, Roelen BAJ, Lawson KA, Zwijsen A. The development of the amnion in mice and other amniotes. Phil Trans R Soc B. 2022;377:20210258.
22. Pereira PN, Dobreva MP, Graham L, Huylebroeck D, Lawson KA, Zwijsen A. Amnion formation in the mouse embryo: the single amniochorionic fold model. BMC Dev Biol. 2011;11:48.
23. Zhu Q, Ge J, Liu Y, Xu J, Yan S, Zhou F. Decoding anterior-posterior axis emergence among mouse, monkey, and human embryos. Dev Cell. 2023;58:63-79.e4.
24. Lawson KA, Meneses JJ, Pedersen RA. Clonal analysis of epiblast fate during germ layer formation in the mouse embryo. Development. 1991;113:891-911.
25. Tam PPL, Beddington RSP. The formation of mesodermal tissues in the mouse embryo during gastrulation and early organogenesis. Development. 1987;99:109-26.
26. O'rahilly R, Müller F. Developmental stages in human embryos. Carnegie Institution Washington, DC; 1987. Available from: https://publicationsonline.carnegiescience.edu/publications_online/developmental_stages.pdf [Last accessed on 3 Jun 2026].
27. O'rahilly R, Müller F. Developmental stages in human embryos: revised and new measurements. Cells Tissues Organs. 2010;192:73-84.
28. Rossant J, Tam PP. New insights into early human development: lessons for stem cell derivation and differentiation. Cell Stem Cell. 2017;20:18-28.
29. Yang H. Spatiotemporal frameworks of morphogenesis and cell lineage specification in pre- and peri-implantation mammalian embryogenesis: insights and knowledge gaps from mouse embryo. Biology. 2025;14:1596.
30. Nakamura T, Fujiwara K, Saitou M, Tsukiyama T. Non-human primates as a model for human development. Stem Cell Rep. 2021;16:1093-103.
31. Schust DJ, Bonney EA, Sugimoto J, et al. The immunology of syncytialized trophoblast. Int J Mol Sci. 2021;22:1767.
32. Gauster M, Moser G, Wernitznig S, Kupper N, Huppertz B. Early human trophoblast development: from morphology to function. Cell Mol Life Sci. 2022;79:345.
33. Kojima J, Ono M, Kuji N, Nishi H. Human chorionic villous differentiation and placental development. Int J Mol Sci. 2022;23:8003.
34. Baergen K, Burton GJ, Baergen RN. Pathology of the human placenta. 6th ed. Berlin, Heidelberg: Springer; 2012. pp. 43-46.
35. Florian J. The formation of the connecting stalk and the extension of the amniotic cavity towards the tissue of the connecting stalk in young human embryos. J Anat. 1930;64:454.
36. Ghimire S, Mantziou V, Moris N, Martinez Arias A. Human gastrulation: the embryo and its models. Dev Biol. 2021;474:100-8.
37. Budjan C, Liu S, Ranga A, Gayen S, Pourquié O, Hormoz S. Paraxial mesoderm organoids model development of human somites. eLife. 2022;11:e68925.
38. Prummel KD, Nieuwenhuize S, Mosimann C. The lateral plate mesoderm. Development. 2020;147:dev175059.
39. Canu G, Ruhrberg C. First blood: the endothelial origins of hematopoietic progenitors. Angiogenesis. 2021;24:199-211.
40. Burton GJ, Jauniaux E. Placentation in the human and higher primates. In: Geisert RD, Spencer T, editors. Placentation in mammals: tribute to EC amoroso’s lifetime contributions to viviparity. Springer; 2021. pp. 223-54.
41. Streeter GL. The Miller ovum: the youngest normal human embryo thus far known. Contrib Embryol Carnegie Inst. 1926;18:31-48. Available from: https://eurekamag.com/research/092/807/092807357.php?__cf_chl_tk=7Y3qKbU7kGQR.X.xnOrPLtgLgIjr5ZC9z8v3.elxhgk-1780312989-1.0.1.1-sXy5TQAKx5BCzvwNx..BlZ4RVbba9PvZFXqn8d4vsf8 [Last accessed on 3 Jun 2026].
42. Boucher D, Pedersen R. Induction and differentiation of extra-embryonic mesoderm in the mouse. Reprod Fertil Dev. 1996;8:765-77.
43. Zhang H, Bradley A. Mice deficient for BMP2 are nonviable and have defects in amnion/chorion and cardiac development. Development. 1996;122:2977-86.
44. Goumans M, Zwijsen A, Rooijen MAV, Huylebroeck D, Roelen BAJ, Mummery CL. Transforming growth factor-β signalling in extraembryonic mesoderm is required for yolk sac vasculogenesis in mice. Development. 1999;126:3473-83.
45. Mallet C, Vittet D, Feige JJ, Bailly S. TGFβ1 induces vasculogenesis and inhibits angiogenic sprouting in an embryonic stem cell differentiation model: respective contribution of ALK1 and ALK5. Stem Cells. 2009;24:2420-7.
46. Carvalho RLC, Itoh F, Goumans M, et al. Compensatory signalling induced in the yolk sac vasculature by deletion of TGFβ receptors in mice. J Cell Sci. 2007;120:4269-77.
47. Firulli AB, Mcfadden DG, Lin Q, Srivastava D, Olson EN. Heart and extra-embryonic mesodermal defects in mouse embryos lacking the bHLH transcription factor Hand1. Nat Genet. 1998;18:266-70.
48. Downs KM. The mouse allantois: new insights at the embryonic-extraembryonic interface. Phil Trans R Soc B. 2022;377:20210251.
49. Nahaboo W, Eski SE, Despin-Guitard E, et al. Keratin filaments mediate the expansion of extra-embryonic membranes in the post-gastrulation mouse embryo. EMBO J. 2022;41:EMBJ2021108747.
50. Hadas R, Rubinstein H, Mittnenzweig M, et al. Temporal BMP4 effects on mouse embryonic and extraembryonic development. Nature. 2024;634:652-61.
51. Rossant J, Cross JC. Placental development: Lessons from mouse mutants. Nat Rev Genet. 2001;2:538-48.
52. Aoki M, Mieda M, Ikeda T, Hamada Y, Nakamura H, Okamoto H. R-spondin3 is required for mouse placental development. Dev Biol. 2007;301:218-26.
53. Sekulovski N, Carleton AE, Lin C, Taniguchi K. Primate amnion development. Development. 2025;152:dev204621.
54. Rossant J, Tam PP. Early human embryonic development: Blastocyst formation to gastrulation. Dev Cell. 2022;57:152-65.
55. Goh I, Botting RA, Rose A, et al. Yolk sac cell atlas reveals multiorgan functions during human early development. Science. 2023;381:eadd7564.
57. Alippe Y, Hatterschide J, Coyne CB, Diamond MS. Innate immune responses to pathogens at the maternal-fetal interface. Nat Rev Immunol. 2025;25:869-84.
58. Enders AC, King BF. Formation and differentiation of extraembryonic mesoderm in the rhesus monkey. Am J Anat. 2005;181:327-40.
59. Hyun I, Bredenoord AL, Briscoe J, Klipstein S, Tan T. Human embryo research beyond the primitive streak. Science. 2021;371:998-1000.
60. Peng Y, Huang X, Zhou Q. Ethical and policy considerations for human embryo and stem cell research in China. Cell Stem Cell. 2020;27:511-4.
61. Farkas K, Ferretti E. Derivation of human extraembryonic mesoderm-like cells from primitive endoderm. Int J Mol Sci. 2023;24:11366.
62. Ai Z, Niu B, Yin Y, et al. Dissecting peri-implantation development using cultured human embryos and embryo-like assembloids. Cell Res. 2023;33:661-78.
63. Hill JP. II. Croonian lecture. - the developmental history of the primates. Philos Trans R Soc Lond B Biol Sci . 1932;221:45-178.
64. Abell AN, Jordan NV, Huang W, et al. MAP3K4/CBP-regulated H2B acetylation controls epithelial-mesenchymal transition in trophoblast stem cells. Cell Stem Cell. 2011;8:525-37.
65. Enders AC, Schlafke S. Differentiation of the blastocyst of the rhesus monkey. Am J Anat. 2005;162:1-21.
66. Knoth M, Larsen JF. Ultrastructure of a human implantation site. Acta Obstet Gynecol Scand. 2011;51:385-93.
67. Boss AL, Chamley LW, James JL. Placental formation in early pregnancy: how is the centre of the placenta made? Hum Reprod Update. 2018;24:750-60.
68. Florian J. The early development of man, with special reference to the development of the mesoderm and cloacal membrane. J Anat. 1933;67:263.
69. Yang R, Goedel A, Kang Y, et al. Amnion signals are essential for mesoderm formation in primates. Nat Commun. 2021;12:5126.
70. Gérard P. Etudes sur l'ovogenese et l'ontogenese chez les Lemuriens du genre Galago. Arch Biol. 1932;43:93-151. Available from: https://eurekamag.com/research/022/591/022591763.php [Last accessed on 3 Jun 2026].
71. Nakamura T, Okamoto I, Sasaki K, et al. A developmental coordinate of pluripotency among mice, monkeys and humans. Nature. 2016;537:57-62.
72. Oldak B, Wildschutz E, Bondarenko V, et al. Complete human day 14 post-implantation embryo models from naive ES cells. Nature. 2023;622:562-73.
73. Niu Y, Sun N, Li C, et al. Dissecting primate early post-implantation development using long-term in vitro embryo culture. Science. 2019;366:eaaw5754.
74. Pham TXA, Panda A, Kagawa H, et al. Modeling human extraembryonic mesoderm cells using naive pluripotent stem cells. Cell Stem Cell. 2022;29:1346-65.e10.
75. Wang SL, Shi GH, Duan K, Yin Y, Li T. Extraembryonic mesoderm cells derived from human embryonic stem cells rely on Wnt pathway activation. Cell Prolif. 2024;58:e13761.
76. Li J, Zhu Q, Cao J, et al. Cynomolgus monkey embryo model captures gastrulation and early pregnancy. Cell Stem Cell. 2023;30:362-77.e7.
77. Haigh T, Chen C, Jones C, Aplin J. Studies of mesenchymal cells from 1st trimester human placenta: expression of cytokeratin outside the trophoblast lineage. Placenta. 1999;20:615-25.
78. Hislop J, Song Q, Keshavarz FK, et al. Modelling post-implantation human development to yolk sac blood emergence. Nature. 2023;626:367-76.
79. Xiao Z, Cui L, Yuan Y, et al. 3D reconstruction of a gastrulating human embryo. Cell. 2024;187:2855-74.e19.
80. Cui L, Lin S, Yang X, et al. Spatial transcriptomic characterization of a Carnegie stage 7 human embryo. Nat Cell Biol. 2025;27:360-9.
81. Gong Y, Bai B, Sun N, et al. Ex utero monkey embryogenesis from blastocyst to early organogenesis. Cell. 2023;186:2092-110.e23.
82. Mendjan S, Mascetti VL, Ortmann D, et al. NANOG and CDX2 pattern distinct subtypes of human mesoderm during exit from pluripotency. Cell Stem Cell. 2014;15:310-25.
83. Richter A, Valdimarsdottir L, Hrafnkelsdottir HE, et al. BMP4 promotes EMT and mesodermal commitment in human embryonic stem cells via SLUG and MSX2. Stem Cells. 2014;32:636-48.
84. Bernardo AS, Faial T, Gardner L, et al. BRACHYURY and CDX2 mediate BMP-induced differentiation of human and mouse pluripotent stem cells into embryonic and extraembryonic lineages. Cell Stem Cell. 2011;9:144-55.
85. Markouli C, De Deckersberg EC, Dziedzicka D, et al. Sustained intrinsic WNT and BMP4 activation impairs hESC differentiation to definitive endoderm and drives the cells towards extra-embryonic mesoderm. Sci Rep. 2021;11:8242.
86. Hu W, Sancho-Serra C, Gantner CW, et al. Atlas of amnion development during the first trimester of human pregnancy. Nat Cell Biol. 2025;27:1175-85.
87. James JL, Lissaman A, Nursalim YNS, Chamley LW. Modelling human placental villous development: designing cultures that reflect anatomy. Cell Mol Life Sci. 2022;79:384.
88. Bárcena A, Muench MO, Kapidzic M, Fisher SJ. A new role for the human placenta as a hematopoietic site throughout gestation. Reprod Sci. 2009;16:178-87.
89. Calvanese V, Mikkola HKA. The genesis of human hematopoietic stem cells. Blood. 2023;142:519-32.
90. Boyd JD, Hamilton WJ. The Human Placenta. Heffer; 1970. Available from: https://books.google.com.sg/books/about/The_human_placenta.html?id=dKtqAAAAMAAJ&redir_esc=y [Last accessed on 3 Jun 2026].
91. Männer J. When does the human embryonic heart start beating? A review of contemporary and historical sources of knowledge about the onset of blood circulation in man. J Cardiovasc Dev Dis. 2022;9:187.
93. Dieterlen-Livre F, Corbel C, Salan J. Allantois and placenta as developmental sources of hematopoietic stem cells. Int J Dev Biol. 2010;54:1079-87.
94. Zeigler BM, Sugiyama D, Chen M, Guo Y, Downs KM, Speck NA. The allantois and chorion, when isolated before circulation or chorio-allantoic fusion, have hematopoietic potential. Development. 2006;133:4183-92.
95. Bulger EA, Mcdevitt TC, Bruneau BG. CDX2 dose-dependently influences the gene regulatory network underlying human extraembryonic mesoderm development. Biology Open. 2024;13:bio060323.
96. Gekas C, Dieterlen-Lièvre F, Orkin SH, Mikkola HK. The placenta is a niche for hematopoietic stem cells. Dev Cell. 2005;8:365-75.
97. Mikkola HK, Gekas C, Orkin SH, Dieterlen-Lievre F. Placenta as a site for hematopoietic stem cell development. Exp Hematol. 2005;33:1048-54.
98. Dancis J, Samuels BD, Douglas GW. Immunological competence of placenta. Science. 1962;136:382-3.
99. Alvarez-Silva M, Belo-Diabangouaya P, Salaün J, Dieterlen-Lièvre F. Mouse placenta is a major hematopoietic organ. Development. 2003;130:5437-44.
100. Azevedo Portilho N, Pelajo-Machado M. Mechanism of hematopoiesis and vasculogenesis in mouse placenta. Placenta. 2018;69:140-5.
101. Demir R, Kayisli U, Seval Y, et al. Sequential expression of VEGF and its receptors in human placental villi during very early pregnancy: differences between placental vasculogenesis and angiogenesis. Placenta. 2004;25:560-72.
102. Lash G, Naruse K, Innes B, Robson S, Searle R, Bulmer J. Secretion of angiogenic growth factors by villous cytotrophoblast and extravillous trophoblast in early human pregnancy. Placenta. 2010;31:545-8.
103. Ivanovs A, Rybtsov S, Ng ES, Stanley EG, Elefanty AG, Medvinsky A. Human haematopoietic stem cell development: from the embryo to the dish. Development. 2017;144:2323-37.
104. Gritz E, Hirschi KK. Specification and function of hemogenic endothelium during embryogenesis. Cell Mol Life Sci. 2016;73:1547-67.
105. Robin C, Bollerot K, Mendes S, et al. Human placenta is a potent hematopoietic niche containing hematopoietic stem and progenitor cells throughout development. Cell Stem Cell. 2009;5:385-95.
106. Calvanese V, Capellera-Garcia S, Ma F, et al. Mapping human haematopoietic stem cells from haemogenic endothelium to birth. Nature. 2022;604:534-40.
107. Reyes L, Golos TG. Hofbauer cells: their role in healthy and complicated pregnancy. Front Immunol. 2018;9:2628.
108. Jiang X, Zhai J, Xiao Z, et al. Identifying a dynamic transcriptomic landscape of the cynomolgus macaque placenta during pregnancy at single-cell resolution. Dev Cell. 2023;58:806-21.e7.
109. Thomas JR, Appios A, Zhao X, et al. Phenotypic and functional characterization of first-trimester human placental macrophages, Hofbauer cells. J Exp Med. 2021;218:e20200891.
110. Thomas JR, Appios A, Calderbank EF, et al. Primitive haematopoiesis in the human placenta gives rise to macrophages with epigenetically silenced HLA-DR. Nat Commun. 2023;14:1764.
111. Van Handel B, Prashad SL, Hassanzadeh-kiabi N, et al. The first trimester human placenta is a site for terminal maturation of primitive erythroid cells. Blood. 2010;116:3321-30.
112. Yao Y, Xu X, Jin L. Macrophage polarization in physiological and pathological pregnancy. Front Immunol. 2019;10:792.
113. Reyes L, Wolfe B, Golos T. Hofbauer cells: placental macrophages of fetal origin. In: Kloc M, editors. Macrophages: origin, functions and biointervention. Cham: Springer; 2017. pp. 45-60.
114. Castellucci M, Celona A, Bartels H, Steininger B, Benedetto V, Kaufmann P. Mitosis of the Hofbauer cell: possible implications for a fetal macrophage. Placenta. 1987;8:65-76.
Cite This Article
How to Cite
Download Citation
Export Citation File:
Type of Import
Tips on Downloading Citation
Citation Manager File Format
Type of Import
Direct Import: When the Direct Import option is selected (the default state), a dialogue box will give you the option to Save or Open the downloaded citation data. Choosing Open will either launch your citation manager or give you a choice of applications with which to use the metadata. The Save option saves the file locally for later use.
Indirect Import: When the Indirect Import option is selected, the metadata is displayed and may be copied and pasted as needed.
About This Article
Special Topic
Copyright

Data & Comments
Data
0













Comments
Comments must be written in English. Spam, offensive content, impersonation, and private information will not be permitted. If any comment is reported and identified as inappropriate content by OAE staff, the comment will be removed without notice. If you have any queries or need any help, please contact us at support@oaepublish.com.