


The potential value of long non-coding RNA in molecular diagnosis and forensic identification of heart failure
 0
0 Abstract
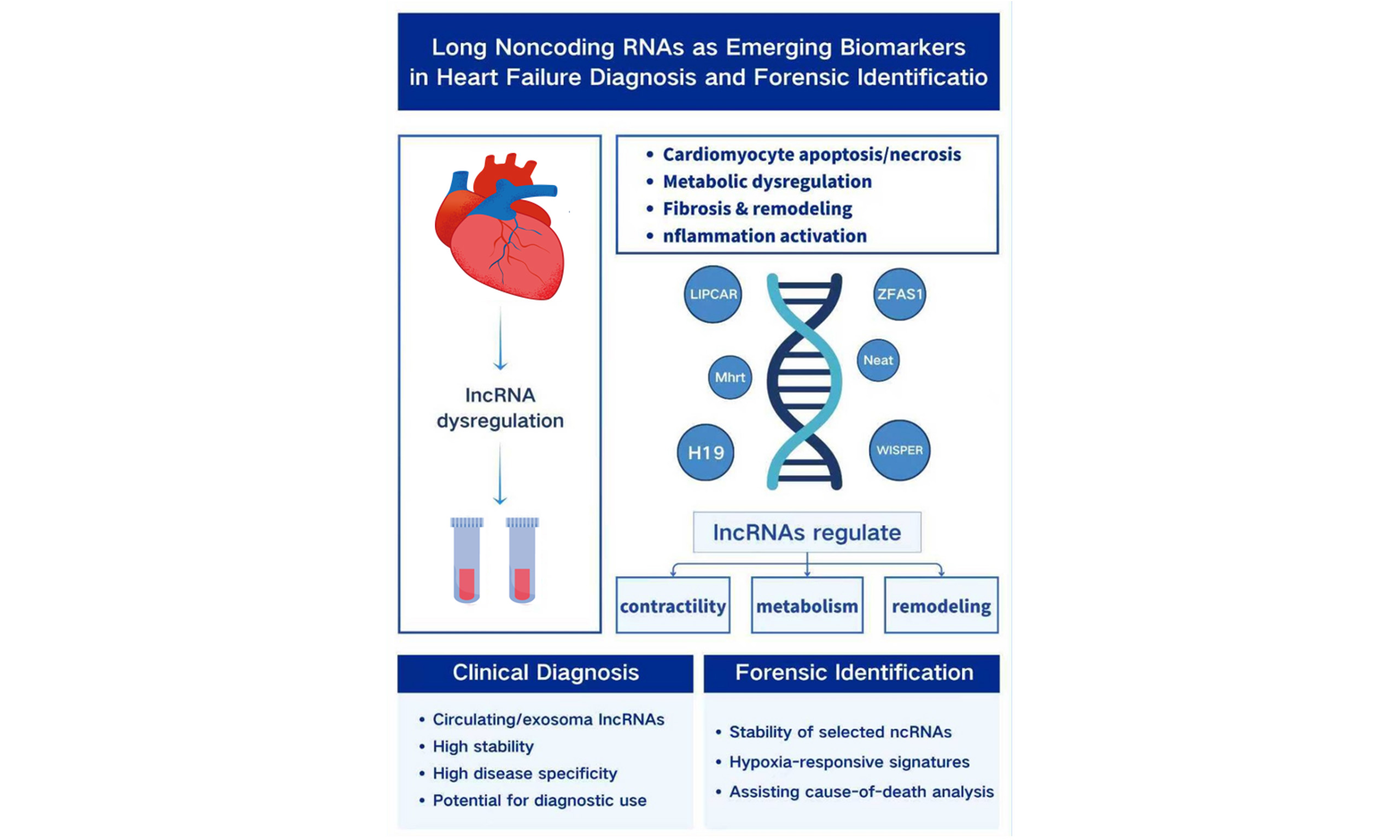
Heart failure (HF), a major cause of cardiovascular mortality worldwide, develops through a multifaceted process involving cardiomyocyte apoptosis and necrosis, metabolic dysregulation, fibrosis, and sustained inflammatory activation. Although B-type natriuretic peptide (BNP) and N-terminal pro-B-type natriuretic peptide (NT-proBNP) are routinely used in clinical assessment, their susceptibility to external influences limits their capacity to fully capture the molecular progression of HF. Emerging evidence has identified long non-coding RNAs (lncRNAs) as important epigenetic regulators that contribute to reduced myocardial contractility and ventricular remodeling. Clinical studies further indicate that circulating lncRNAs - particularly those packaged within exosomes - possess notable stability and strong disease specificity, making them promising candidates for improving diagnostic and prognostic evaluation. With ongoing advances in transcriptomic profiling and exosome-based technologies, the potential use of lncRNAs is also extending into forensic identification. This review synthesizes current insights into lncRNA-mediated regulation in HF progression, evaluates recent developments in circulating lncRNA detection, and explores their emerging forensic applications. It also outlines key limitations in existing research and discusses future directions to support deeper investigation into the molecular mechanisms underlying cardiac death and their relevance to forensic practice.
Keywords
INTRODUCTION
Heart failure (HF) is a clinical syndrome characterized by structural and/or functional cardiac abnormalities that emerge during the decompensated stage of progressive cardiac dysfunction. It is commonly associated with elevated levels of natriuretic peptides and objective evidence of pulmonary and/or systemic congestion. As the end stage of a wide range of cardiovascular diseases, HF carries substantial mortality, most commonly due to sudden cardiac death (accounting for more than half of all cases) or from multiorgan dysfunction resulting from inadequate perfusion[1-3]. Impairment of myocardial systolic or diastolic function reduces cardiac output, rendering the heart unable to meet the body’s demands for oxygen and nutrients. As cardiac function progressively deteriorates, vital organs (such as the kidneys, liver, and brain) develop dysfunction or failure due to decreased perfusion and/or venous congestion[4]. Recent epidemiological updates estimate that more than 64 million people worldwide are living with heart failure, and the five-year mortality rate remains close to 50%. Although the incidence of HF has shown a modest decline, its overall prevalence continues to rise, largely owing to advances in medical care and increased life expectancy. At the same time, despite a 7% reduction in age-standardized prevalence, substantial differences persist across countries. The absolute number of individuals with HF has increased by 92%, and disability-adjusted life years have risen by 106%, reflecting the growing global burden of the disease[5,6]. Levels of B-type natriuretic peptide (BNP) and N-terminal pro-B-type natriuretic peptide (NT-proBNP) are elevated in heart failure, with their concentrations rising in proportion to the severity and duration of ventricular dysfunction. These peptides serve as established biomarkers of HF and are widely regarded as the gold standard for prognostic evaluation and risk stratification. However, their diagnostic sensitivity and specificity are reduced in patients with diastolic dysfunction or in those presenting with acute decompensated heart failure[7],posing significant challenges for forensic pathologists determining the manner of death. Long non-coding RNA (lncRNA) has emerged as a key participant in cardiac function, demonstrating potential as a therapeutic target for HF[8]. Long non-coding RNAs (lncRNAs) are RNA transcripts longer than 200 nucleotides that lack protein-coding capacity[9]. Once considered transcriptional noise, they are now recognized as key regulators of gene transcription, translation, and epigenetic modification, participating in essential pathways that govern cardiovascular function[8]. lncRNAs play pivotal roles in both physiological and pathological cardiac processes and have emerged as promising therapeutic targets for heart failure. They significantly influence myocardial fibrosis and the progression of heart failure following myocardial infarction[10], contributing to the regulation of myocardial contractility, cardiac hypertrophy, cardiomyocyte apoptosis, and fibrotic remodeling. To better illustrate the relative advantages and limitations of long non-coding RNAs, we provide a comparison between lncRNAs and other commonly used molecular biomarkers for heart failure is [Table 1].
Comparison of lncRNA and other molecular biomarkers for heart failure diagnosis and forensic application
| Biomarker type | Main advantages | Main limitations |
| lncRNA | High tissue and disease specificity and involved in key regulatory pathways of myocardial remodeling and fibrosis[11] Detectable in blood tissues[8] Relatively stable when packaged in extracellular vesicles[12] | Limited standardization of detection methods[8] Functional roles of many lncRNAs remain incompletely characterized[8] Lack of large-scale clinical and forensic validation[13] |
| miRNA | High stability in circulation[14] Extensively studied in cardiovascular diseases[5] | Lower tissue specificity and often reflects general stress or injury responses[14] Limited accuracy and poor reproducibility as standalone biomarkers[15] |
| Protein biomarkers (e.g., BNP, NT-proBNP) | Widely used in clinical practice[5] Standardized assays[16] Strong association with HF severity[5] | Affected by renal function, age, and obesity[17] Limited applicability in postmortem samples due to degradation[1] |
Notably, non-coding RNAs particularly lncRNAs can be selectively packaged into extracellular vesicles (EVs), through which they modulate pathophysiological processes in recipient cells, making them attractive candidates for heart failure diagnosis and therapy[13]. As stable RNA carriers, EVs preserve lncRNA integrity in circulating fluids and postmortem samples, offering new possibilities for molecular investigations of cardiac death[18]. Through their ability to regulate specific proteins and biological pathways, lncRNAs hold significant therapeutic potential in heart failure, enabling targeted intervention at the level of RNA function[18]. Their unique biological characteristics and detectability in circulation also suggest potential value in forensic applications. However, studies examining the diagnostic and forensic relevance of lncRNAs in heart failure remain limited. This review therefore aims to explore the regulatory mechanisms of lncRNAs during the decompensated phase of heart failure and assess their potential forensic applications, with the goal of providing new insights for future forensic research. Although numerous reviews have addressed the role of lncRNAs in cardiovascular disease and heart failure, most have focused on their molecular mechanisms or therapeutic implications in living patients. In contrast, the diagnostic challenges associated with the decompensated phase of HF and the potential forensic relevance of lncRNAs have received little systematic attention. This review specifically integrates current knowledge of lncRNA-mediated regulatory pathways in decompensated HF with emerging evidence from postmortem and extracellular vesicle research, thereby highlighting their potential utility as molecular markers for forensic diagnosis and cause-of-death determination.
EXPLORING THE MECHANISMS OF LNCRNA IN HEART FAILURE
The precise mechanisms underlying HF remain incompletely understood despite extensive clinical and experimental investigation[8]. Data from the European Society of Cardiology (ESC) Long-Term Registry indicate that among outpatients with diagnosed HF, approximately 60% presented with heart failure with reduced ejection fraction (HFrEF), 24% with heart failure with mildly reduced ejection fraction (HFmrEF), and 16% with heart failure with preserved ejection fraction (HFpEF)[17]. These epidemiological insights highlight the importance of focusing on molecular mechanisms associated with reduced ejection fraction to deepen our understanding of HF pathogenesis.The diagnostic criteria for HFrEF include a left ventricular ejection fraction (LVEF) ≤ 40%, indicating significant impairment of systolic function. In contrast, the clinical diagnostic criteria for HFpEF include a preserved (or near-normal) LVEF, typically defined as LVEF ≥ 50%. For both conditions, supportive diagnostic criteria require the presence of symptoms and/or signs of HF, such as dyspnea, fatigue, peripheral edema, pulmonary congestion, jugular venous distention, hepatojugular reflux, and a third heart sound (S3)[19].
With the advancement of high-throughput transcriptomic technologies, lncRNAs have emerged as important regulatory molecules in cardiovascular biology. Over the past decade, increasing evidence has demonstrated that lncRNAs participate in key pathological processes of HF, including cardiomyocyte hypertrophy, apoptosis, fibrosis, inflammation, and calcium handling[8]. Specific lncRNAs have been shown to modulate gene expression at transcriptional, post-transcriptional, and epigenetic levels, thereby influencing ventricular contractility and structural remodeling[20]. These findings highlight the critical role of lncRNAs as upstream regulators within complex molecular networks underlying HF progression.From a forensic perspective, determining the cause of death often requires integrating molecular alterations with histomorphological observations. Studying how changes in myocardial contractility translate into structural and functional remodeling during the decompensated phases of HF can therefore provide critical insights for medicolegal evaluations[1,8]. Consequently, summarizing the roles of lncRNAs in regulating ventricular contractile function is valuable for guiding future mechanistic investigations and assisting pathologists in interpreting postmortem cardiac alterations [Table 2].
lncRNAs associated with reduced myocardial contractility
| lncRNA | Sample source | Mechanism axis | Effect on HF progression | Authors |
| Mhrt | Mouse Model and Cardiomyocytes | Mhrt - Brg1 - stress gene promoters | Inhibits HF progression | Han et al., 2014[21] |
| NEAT1 | AMI model mice and HL-1 hypoxic cells | NEAT1 - miR-450b-5p - lipid peroxidation | Promotes HF progression | Ge et al., 2022[22] Yu et al., 2024[23] |
| LIPCAR | Plasma/exosome | LIPCAR - mitochondrial dysfunction | Promotes HF progression | Yan et al., 2021[24] Turkieh et al.[25] |
| MIAT | Cardiac hypertrophy models | MIAT - YTHDF2 - PPARα/CPT-1a | Promotes HF progression | Yang et al.[26] |
| ZFAS1 | Myocardium in mouse myocardial infarction model | ZFAS1 - SERCA2a - Ca2+ overload | Promotes HF progression | Zhang et al., 2018[27] |
| H19 | Mouse myocardial infarction and hypoxia model | H19 - miR-675 - VDAC1; H19 - YB-1 - COL1A1 | Bidirectional effect on HF progression | Su et al., 2021[28] Choong et al.[29] |
| UCA1 | TAC mouse myocardial tissue | UCA1 - miR-184 - HOXA9 | Promotes HF progression | Li et al., 2021[30] |
| KCNQ1OT1 | OGD/R-induced H9c2 injury model | KCNQ1OT1 - AdipoR1 - p38/NF-κB | Promotes HF progression | Li et al.[31] |
Changes in myocardial contractile proteins
A pivotal moment in lncRNA research for HF was the demonstration that Mhrt can prevent pathological myocardial hypertrophy by antagonizing Brg1 (brahma-related gene 1), establishing a direct link between lncRNAs and epigenetic regulation of cardiac contractile proteins.
Mhrt (myosin heavy chain-related RNA transcript) was one of the first cardiac-enriched lncRNAs identified to play a functional role in pathological cardiac remodeling. It was initially discovered through transcriptomic profiling of stressed murine hearts, where a cluster of previously unannotated long non-coding RNAs was found to be transcribed from the Myh7 locus. Among these, Mhrt was shown to be highly and specifically expressed in cardiomyocytes and dynamically regulated under mechanical stress, suggesting its involvement in cardiac contractile regulation. Mechanistically, Mhrt directly binds to Brg1, a core component of the chromatin remodeling complex, thereby blocking Brg1 from interacting with stress-responsive promoter regions. This action suppresses transcriptional reprogramming induced by mechanical stress. Under physiological conditions, Mhrt maintains chromatin stability and transcriptional homeostasis in cardiomyocytes. However, under sustained mechanical stress or pathological conditions, Mhrt expression is markedly downregulated, allowing Brg1 hyperactivation. This dysregulation triggers abnormal expression of genes associated with myocardial hypertrophy, fibrosis, and metabolic imbalance, ultimately accelerating ventricular remodeling and the progression to HF[21].
Advances in transcriptomics and functional analyses have since identified numerous lncRNAs - including NEAT1 (nuclear enriched abundant transcript 1), H19 (H19 imprinted maternally expressed transcript), WISPER (Wisp2 super-enhancer-associated RNA), ZFAS1 (Zinc Finger Antisense 1), and LIPCAR (Long Intergenic non-coding RNA predicting CARdiac remodeling) - that are implicated in hypertrophy, fibrosis, metabolic reprogramming, and cardiomyocyte death. The breadth and pace of experimental and clinical research on lncRNAs have expanded rapidly in recent years, reflecting their central role in cardiac pathophysiology[3].
Cardiac muscle contraction primarily depends on contractile proteins[32], with cardiomyocytes and cardiac fibroblasts playing essential roles in generating contractile force and synthesizing structural proteins[33]. In pathological conditions such as myocardial fibrosis and hypertrophy, apoptosis and necrosis of cardiomyocytes reduce the number of functional cells, impair protein synthesis, and compromise overall myocardial contractile function. NEAT1 has been demonstrated to exert regulatory effects on fibrosis across multiple organs, including the heart, lung, liver, and kidney[34]. NEAT1 influences the pathophysiology of myocardial infarction, HF, and atherosclerosis by modulating inflammation, apoptosis, and oxidative stress, processes closely associated with cardiomyocyte ferroptosis[22]. Mechanistically, NEAT1 binds directly to miR-450b-5p, negatively regulating its expression. NEAT1 expression is markedly upregulated in hypoxic HL-1 mouse atrial cardiomyocyte cells and in acute myocardial infarction (AMI) mouse models, whereas NEAT1 silencing alleviates lipid peroxidation and mitigates myocardial ferroptosis both in vitro and in vivo[23].
At present, increasing attention has been directed toward myocardial injury-induced impairment of cardiac contractile function. Aberrant overexpression of Myocardial Infarction-Associated Transcript (MIAT) significantly promotes the initiation and progression of cardiac hypertrophy. Mechanistically, MIAT upregulation is accompanied by enhanced expression of the m6A RNA methylation reader protein YTHDF2, which accelerates suppression of the PPARα/CPT-1a (peroxisome proliferator-activated receptor-α/ carnitine palmitoyltransferase-1a) signaling pathway, leading to reduced stability and decreased expression of carnitine palmitoyltransferase 1a (CPT-1a) messenger RNA (mRNA). This cascade disrupts myocardial energy metabolism and exacerbates adverse structural and functional cardiac remodeling, indicating that MIAT may play a deleterious role in pathological cardiac remodeling during the development of myocardial hypertrophy[26]. In myocardial injury models, KCNQ1OT1 is markedly upregulated, and its aberrant expression is closely associated with enhanced inflammatory responses and programmed cell death in cardiomyocytes. Functional studies have demonstrated that suppression of KCNQ1OT1 significantly improves cardiomyocyte viability, reduces apoptosis, and downregulates the expression of pro-inflammatory mediators, including tumor necrosis factor-alpha (TNF-α), interleukin-6 (IL-6), and interleukin-1 beta (IL-1β). Mechanistically, KCNQ1OT1 modulates the activation status of the p38 mitogen-activated protein kinase/nuclear factor kappa B (MAPK/NF-κB) signaling pathway by regulating the adiponectin receptor AdipoR1 (adiponectin receptor 1), thereby influencing the inflammation-apoptosis signaling network in cardiomyocytes. Aberrant activation of this regulatory axis exacerbates cardiomyocyte injury and functional impairment, suggesting that KCNQ1OT1-mediated signaling pathways may contribute to the molecular basis of heart failure progression and represent promising targets for diagnostic and therapeutic intervention[31]. H9c2 cells are a rat embryonic ventricular-derived cardiac cell line that is widely used to model the biological responses of cardiomyocytes under various pathological conditions. The long non-coding RNA ANRIL (antisense non-coding RNA in the INK4 locus) is significantly upregulated in hypoxia-induced cardiomyocyte injury models and plays an important regulatory role in cytoprotective mechanisms. Functional experiments have shown that silencing ANRIL markedly aggravates hypoxia-induced injury in H9c2 cardiomyocytes, as evidenced by reduced cell viability, increased apoptosis, and enhanced oxidative stress.Mechanistic studies have demonstrated that ANRIL acts as a competing endogenous RNA (ceRNA) by specifically binding to miR-7-5p, thereby relieving its inhibitory effect on the downstream target gene SIRT1 and promoting SIRT1 expression. Further investigations revealed that inhibition of miR-7-5p or overexpression of SIRT1 significantly attenuates the exacerbated hypoxia-induced cellular injury caused by ANRIL deficiency. Collectively, these findings suggest that the ANRIL-miR-7-5p-SIRT1 regulatory axis exerts a critical protective effect in hypoxia-associated cardiomyocyte damage and may participate in the molecular regulatory network underlying the development and progression of heart failure[35].
Myocardial energy metabolism dysfunction
Metabolic disturbances represent a central pathological mechanism in HF. The heart, as an organ with exceptionally high oxygen demand, relies predominantly on mitochondrial oxidative phosphorylation to generate adenosine triphosphate (ATP). In humans, approximately 65 kilograms of ATP are synthesized and consumed daily, with the heart accounting for roughly 8% of this total[36]. Beyond ATP production, mitochondria are essential for cardiomyocyte metabolism, calcium (Ca2+) homeostasis, lipid synthesis, and redox regulation. Investigating the expression and function of mitochondrial-associated lncRNAs in HF can provide deeper insights into mitochondrial dysfunction during disease progression[37]. Notably, the mitochondrial-associated lncRNA LIPCAR is significantly elevated in the plasma of patients with HF following AMI. Its plasma levels correlate closely with left ventricular remodeling and adverse clinical outcomes. Receiver operating characteristic (ROC) analysis indicates that LIPCAR holds potential as an early diagnostic biomarker for HF[24]. In addition to LIPCAR, other cytoplasmic lncRNAs have also attracted increasing attention. CHKB divergent transcript (CHKB-DT) is a cardiac-enriched lncRNA that predominantly localizes in the cytoplasm and maintains mitochondrial metabolic homeostasis by stabilizing ALDH2 mRNA through interaction with the RNA-binding protein fused in sarcoma (FUS). Loss of CHKB-DT leads to accumulation of 4-hydroxy-2-nonenal, mitochondrial dysfunction, and impaired cardiomyocyte contractility, thereby promoting the development and progression of dilated cardiomyopathy. These findings indicate that CHKB-DT plays a critical regulatory role in ventricular remodeling[38]. Cardiomyocyte excitation represents an electrical process, whereas myofilament contraction constitutes a mechanical response. Ca2+ plays a pivotal role in transducing electrical signals into mechanical contraction. Disruption of Ca2+ transport or intracellular distribution can impair excitation-contraction coupling, thereby compromising both contraction and relaxation of cardiomyocytes[39]. Interaction between ZFAS1 and SERCA2a (serine/threonine-dependent calcium ATPase 2a, a key protein in the sarcoplasmic reticulum membrane) leads to intracellular calcium overload. This dysregulation impairs the contractile capacity of cardiomyocytes, contributing to diminished myocardial function[27].
Ventricular remodeling
Ventricular remodeling is a chronic and multifaceted adaptive response to myocardial injury or increased hemodynamic load. It involves extensive alterations in gene expression across multiple signaling pathways, resulting in structural, metabolic, and functional changes in the ventricle. This time-dependent process is relevant not only to HF progression but also to forensic analyses, where it can inform determinations of cause of death in chronic cardiac conditions[1,40,41].
Emerging evidence underscores the role of lncRNAs as key modulators of ventricular remodeling. For example, in hypertrophic cardiomyocytes, lncRNA UCA1 regulates remodeling via the miR-184/HOXA9 axis. In mouse models subjected to transverse aortic constriction (TAC), UCA1 expression is significantly upregulated, whereas miR-184 is downregulated[30]. Similarly, lncRNA H19 exhibits dual regulatory effects during myocardial hypertrophy. H19, a maternally imprinted gene with high spatiotemporal specificity, mediates cell-type-specific responses through multiple signaling pathways[28,29]. Under mechanical stretch or increased pressure load, H19 expression is elevated, which inhibits the abnormal opening of voltage-dependent anion channel 1 (VDAC1) via the H19/miR-675/VDAC1 axis, thereby reducing cellular energy stress and apoptosis, exerting a compensatory protective effect[42]. Conversely, under hypoxic conditions, H19 interacts with YB-1 (Y-box binding protein-1) to suppress Collagen1A1 expression, promoting fibrosis. In mouse models of ischemic injury, H19 is markedly upregulated in infarcted regions alongside multiple extracellular matrix (ECM) genes. CRISPR-Cas9-mediated knockout of H19 improves remodeling outcomes and reduces ECM gene expression, demonstrating that H19 overexpression contributes to pathological cardiac dilation and fibrosis[29]. Chaer is a cardiac-enriched epigenetic lncRNA that regulates ventricular remodeling through stress-induced interaction with the PRC2 complex. Upon hypertrophic stimulation, Chaer transiently binds PRC2 in an mTORC1-dependent manner, disrupting its genomic targeting and reducing H3K27me3 at hypertrophy-associated loci. This epigenetic switch promotes pathological gene reprogramming, leading to cardiomyocyte hypertrophy and fibrotic remodeling. Genetic inactivation of Chaer attenuates stress-induced hypertrophy and fibrosis, highlighting its critical role in early-stage ventricular remodeling[43]. Chast(cardiac hypertrophy-associated transcript) is a lncRNA specifically upregulated in cardiomyocytes during pressure overload-induced hypertrophy and in human hypertrophic heart tissue. Experimental overexpression of Chast is sufficient to induce cardiomyocyte hypertrophy, whereas its silencing attenuates pathological cardiac remodeling. Mechanistically, Chast promotes hypertrophy by repressing Pleckstrin homology domain-containing protein family M member 1 and impairing cardiomyocyte autophagy, indicating its critical role in regulating cardiac hypertrophy and ventricular remodeling[44].
Collectively, these findings highlight the dual role of lncRNAs in HF progression and illustrate that distinct lncRNAs may influence cardiovascular disease pathogenesis through shared or convergent signaling pathways, underscoring their potential as therapeutic targets and biomarkers.
Translation of lncRNAs into micropeptides
Although lncRNAs were originally defined as non-coding transcripts, recent studies have shown that a subset of lncRNAs contains actively translated small open reading frames that generate functional micropeptides. Ribosome profiling in human heart tissue revealed that approximately 20% of expressed lncRNAs undergo translation.Several lncRNAs previously implicated in cardiac remodeling, including NEAT1, ZFAS1, Hand2-associated long non-coding RNA UPPERHAND, and chaer, were found to produce micropeptides, indicating that lncRNA loci may exert biological effects through both RNA- and peptide-mediated mechanisms[45]. These micropeptides are frequently associated with mitochondrial and membrane-related pathways and exhibit coordinated expression with genes involved in oxidative phosphorylation and cardiac metabolism. Moreover, multiple translated lncRNAs are differentially expressed in hypertrophic and failing hearts, suggesting their involvement in stress-induced myocardial remodeling. Collectively, these findings expand the functional repertoire of lncRNAs and highlight lncRNA-derived micropeptides as a novel regulatory layer in HF pathogenesis and a potential source of diagnostic biomarkers and therapeutic targets[46].
Collectively, these findings demonstrate that lncRNAs influence cardiac function through coordinated regulation of contractile machinery, metabolic homeostasis, calcium handling, and structural remodeling, thereby contributing to impaired myocardial performance and reduced ejection capacity [Figure 1].
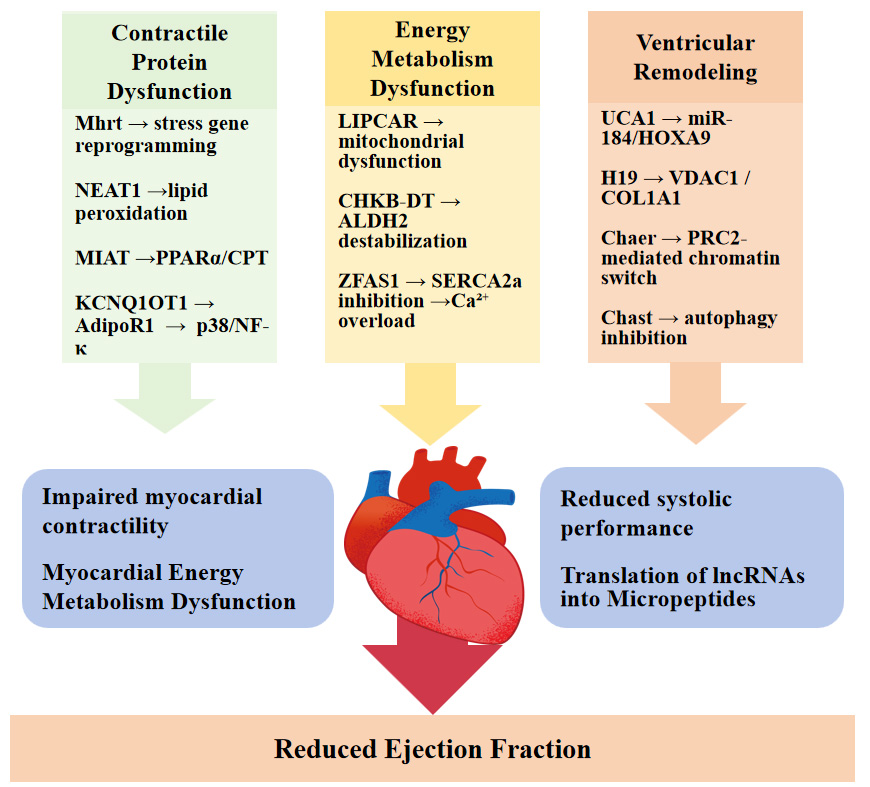
Figure 1. Mechanistic roles of lncRNAs in the regulation of myocardial contractility. Schematic illustration of the major molecular pathways through which long non-coding RNAs (lncRNAs) participate in heart failure-related myocardial dysfunction. lncRNAs regulate cardiac pathology at multiple levels, including myocardial contractile protein expression, energy metabolism, ventricular remodeling, and translation into functional micropeptides. Through modulation of key signaling axes, these regulatory processes collectively promote cardiomyocyte injury, fibrosis, metabolic imbalance, and calcium handling disorders, ultimately leading to reduced cardiac ejection capacity. NEAT1: Nuclear enriched abundant transcript 1; MIAT: myocardial infarction-associated transcript; PPARα: peroxisome proliferator-activated receptor alpha; CPT: carnitine palmitoyltransferase; KCNQ1OT1: KCNQ1 opposite strand/antisense transcript 1; NF-κB: nuclear factor kappa B.
DIAGNOSTIC AND PROGNOSTIC VALUE OF LNCRNAS
Limitations of conventional biomarkers in HF diagnosis
Despite substantial advances in clinical medicine, the early diagnosis and accurate prognostic assessment of HF continue to pose significant challenges for clinicians and researchers alike[5]. Conventional biomarkers, including BNP and NT-proBNP, remain widely utilized in routine clinical practice. However, their circulating levels are often influenced by a variety of confounding factors, such as patient age, renal function, fluid status, and ongoing pharmacological treatments. Moreover, the relatively short half-life of these peptides limits their ability to reflect real-time or dynamic molecular changes that occur during the progressive remodeling of cardiac tissue in chronic heart failure. In this context, advances in transcriptomic technologies have propelled lncRNAs to the forefront of HF biomarker research. lncRNAs exhibit tissue-specific expression patterns, remarkable stability in biological fluids, and reproducibility in detection, making them particularly attractive candidates for non-invasive diagnostics[13].
Circulating and exosomal lncRNAs as emerging diagnostic biomarkers
Increasing evidence indicates that circulating or exosome-encapsulated lncRNAs reflect key pathological processes - including cardiomyocyte injury, metabolic imbalance, and fibrotic remodeling - thereby providing promising tools for personalized diagnosis and risk stratification in heart failure. Notably, early landmark studies in patients with AMI established the clinical feasibility of detecting circulating lncRNAs in plasma, laying the methodological foundation for subsequent heart failure-oriented investigations rather than representing disease-specific mechanistic evidence. For example, Turkieh et al. reported that plasma-derived exosomes from AMI patients displayed significantly elevated LIPCAR levels. Importantly, LIPCAR expression showed a negative correlation with LVEF and a positive correlation with left ventricular end-diastolic volume, indicating its close association with adverse ventricular remodeling and providing critical conceptual support for its later application as a biomarker in HF[25].
In subsequent clinical studies focusing on post-AMI heart failure, LIPCAR demonstrated excellent diagnostic performance, with an area under the curve (AUC) of 0.985 for identifying heart failure after AMI, which was markedly higher than that of the conventional biomarker NT-proBNP (AUC = 0.714). This finding suggests that LIPCAR may provide superior sensitivity in reflecting early molecular alterations associated with ventricular remodeling compared with traditional peptide-based markers[24]. Beyond LIPCAR, additional lncRNAs have been implicated in cardiovascular pathology.In patients with acute coronary syndromes, metastasis-associated lung adenocarcinoma transcript 1 (MALAT1) and LIPCAR were also shown to discriminate vulnerable coronary artery disease, with AUC values of 0.773 and 0.737, respectively, while a combined multi-RNA model incorporating LIPCAR, MALAT1, and miR-155-5p further improved predictive performance (AUC = 0.938). These values are comparable to or exceed those reported for several conventional cardiac enzymes in similar clinical settings, highlighting the potential additive value of lncRNAs to established biomarker panels[47].
These observations collectively support the utility of circulating lncRNAs not only as early indicators of post-injury remodeling but also as tools for risk stratification and long-term prognostic evaluation. Complementary evidence comes from transcriptomic analyses in peripheral blood mononuclear cells (PBMCs). Qi et al. identified 1,429 differentially expressed lncRNAs in PBMCs from maintenance hemodialysis patients. Among these, ENST00000561762 exhibited significant correlations with LVEF, proBNP levels, and 6 min walk distance, indicating that lncRNA expression profiles in peripheral blood may serve as reliable proxies for both cardiac function and systemic metabolic status[48].
Current challenges and future perspectives
Collectively, circulating lncRNAs represent a promising new generation of biomarkers for HF. Their high stability, disease-specific expression patterns, and correlation with structural and functional parameters position them as valuable complements to existing protein-based biomarkers. However, large-scale, multicenter prospective studies are still required to validate their clinical utility, standardize detection methodologies, and define threshold values for clinical decision-making. Future research directions include the development of multi-lncRNA diagnostic panels, integration with established biomarkers such as NT-proBNP, and application of machine learning algorithms to improve predictive accuracy. With continued technological and clinical advances, circulating lncRNAs are expected to play an increasingly important role in precision diagnosis and prognostic evaluation of HF.
EXTRACELLULAR VESICLE-DERIVED LNCRNAS IN HEART FAILURE
EVs are membrane-bound nanoscale particles actively released by cells into the extracellular environment and are widely present in blood, urine, saliva, and other body fluids. According to their biogenesis pathways and size ranges, EVs are generally classified into exosomes, microvesicles, and apoptotic bodies[49]. Exosomes originate from the fusion of multivesicular bodies with the plasma membrane, whereas microvesicles are formed through direct outward budding of the plasma membrane[50]. EVs encapsulate a variety of bioactive molecules, including proteins, lipids, and diverse types of nucleic acids, such as microRNAs (miRNAs), lncRNAs, circular RNAs (circRNAs), and DNA fragments[13]. These molecular cargos can be delivered to recipient cells via EVs, thereby mediating intercellular communication and participating in multiple physiological and pathological processes, including inflammatory responses, immune regulation, maintenance of metabolic homeostasis, and tissue remodeling. In recent years, EVs have been recognized as critical signaling carriers linking local tissue injury to systemic responses and are increasingly implicated in the initiation and progression of cardiovascular diseases[51].
Landscape of lncRNAs in extracellular vesicles
In recent years, it has been demonstrated that EVs carry a variety of lncRNAs, whose abundance within EVs often differs from that in their parental cells, suggesting the existence of selective sorting mechanisms. Certain lncRNAs can be preferentially packaged into EVs under specific stress conditions, such as DNA damage or pathological stimuli, indicating that their incorporation is condition-dependent and tightly regulated[52]. Furthermore, some EV-associated lncRNAs harbor sequences or structural motifs capable of interacting with miRNAs or RNA-binding proteins (RBPs), potentially modulating miRNA activity via a “sponge” effect or facilitating targeted miRNA loading, thereby contributing to long-range intercellular communication[53].
Exosomal lncRNAs as functional regulators in heart failure
Recent studies have demonstrated that exosome-derived lncRNAs are closely associated with the development and progression of heart failure and can reflect disease severity as well as therapeutic responses. For instance, exosomal lncRNA KLF3-AS1 is significantly upregulated in patients with HFrEF and shows a positive correlation with NT-proBNP and a negative correlation with left ventricular ejection fraction, with good diagnostic performance (AUC = 0.861). Moreover, KLF3-AS1 suppresses hypertrophic marker expression in cardiomyocyte hypertrophy models, suggesting that exosomal lncRNAs may serve not only as circulating biomarkers but also as functional regulators involved in myocardial remodeling[54]. Meanwhile, accumulating evidence indicates that the plasma exosomal lncRNA expression profile is systemically altered in patients with cardiovascular diseases. For instance, exo-lncR-NEAT1, exo-lncR-HCG15, and exo-lncR-UCA1 are significantly upregulated in affected individuals, highlighting their potential diagnostic value[55]. Collectively, these findings suggest that exosomal lncRNAs not only mirror disease status but may also constitute an important molecular basis for precision diagnosis and therapeutic intervention in cardiovascular diseases.
Although exosome (EV)-derived lncRNAs have shown considerable potential in the early diagnosis and intervention of heart failure and other cardiovascular diseases, their clinical translation remains confronted with multiple challenges. On the one hand, the biological functions of EV-associated lncRNAs exhibit marked heterogeneity depending on cellular origin and distinct physiological or pathological conditions, and their sorting mechanisms and targeting properties have yet to be fully elucidated. On the other hand, there is still a lack of standardized protocols for EV isolation and purification, quantitative normalization, stability assessment, and delivery efficiency. Moreover, substantial variability in biological materials, experimental conditions, and methodological standards across studies greatly limits the generalizability and applicability of current findings. Future investigations should therefore focus on clarifying the underlying molecular mechanisms and further optimizing the targeting capability and biosafety of engineered EVs, thereby facilitating the translation of EV-lncRNAs from experimental research into precision diagnostic and therapeutic applications[51,56].
PROSPECTS OF LNCRNAS FOR CARDIOVASCULAR THERAPY
As an emerging precision medicine strategy, RNA therapy has achieved breakthrough progress in recent years in the fields of genetic diseases, metabolic disorders, and oncology, providing a new theoretical foundation for molecular intervention in cardiovascular diseases[57]. Compared with traditional small-molecule drugs and protein-based therapeutics, RNA therapy offers distinct advantages including high target specificity, well-defined mechanisms of action, and direct modulation of gene expression levels. It is particularly effective for pathogenic genes or regulatory non-coding RNAs that are difficult to target with conventional drug interventions[58]. Currently approved RNA therapeutics primarily include antisense oligonucleotides (e.g., nusinersen and inotersen), small interfering RNAs (e.g., patisiran and inclisiran), and mRNA-based drugs [e.g., COVID-19 vaccines and vascular endothelial growth factor A (VEGF-A) mRNA for ischemic heart disease]. These successful clinical applications have provided important proof-of-concept for the feasibility of RNA-based therapies in the cardiovascular field. Multiple lncRNAs involved in cardiac remodeling exhibit well-defined pathogenic or cardioprotective functions, relatively tissue-specific expression patterns, and molecular properties that allow effective modulation by exogenous interventions, suggesting their potential as therapeutic targets. For instance, inhibition of MALAT1 attenuates endothelial inflammatory responses[59], whereas silencing of Chast alleviates myocardial hypertrophy and cardiac fibrosis[44]. In contrast, cardioprotective lncRNAs such as Mhrt exert anti-heart failure effects in animal models when their expression is enhanced via viral vector-mediated delivery[21]. Collectively, these functional intervention studies provide further support for the feasibility of lncRNAs as potential therapeutic targets for heart failure. Nevertheless, lncRNA-based therapies still face several challenges, including insufficient delivery efficiency, limited tissue specificity, uncertain long-term safety, and potential off-target effects[60]. Overall, with continued optimization of delivery systems and advances in targeting strategies, lncRNAs are expected to be integrated with existing RNA therapeutic platforms and may emerge as an important complementary approach for precision treatment of heart failure.
FORENSIC APPLICATIONS OF LNCRNAS IN HEART FAILURE
Non-coding RNAs (ncRNAs) have emerged as critical molecular tools in various forensic applications, including postmortem interval (PMI) determination, wound age estimation, forensic age assessment, cause-of-death analysis, and bodily fluid identification[61]. Accordingly, forensic researchers have increasingly focused on the role of lncRNAs in postmortem detection and auxiliary identification. The first documented forensic application of ncRNAs dates back to 2009, when Hanson and colleagues investigated the potential utility of miRNAs for bodily fluid identification[62]. Over the past decade, a growing number of studies have explored the applicability of ncRNAs in forensic science[61]. However, translating these findings into routine practice remains challenging due to limited clinical applications and a scarcity of well-validated biomarkers.In the context of heart failure, forensic application of lncRNAs is gaining attention as a potential molecular tool for postmortem evaluation of chronic cardiac remodeling and functional decompensation, particularly in cases with ambiguous structural findings[Table 3].
ncRNA in postmortem circulatory detection
| ncRNA | Sample sources and postmortem characteristics | Forensic Application Value | Authors |
| miR-1-2 | Myocardial tissue Remains relatively stable 8 days postmortem | Demonstrates postmortem stability of ncRNA in myocardial tissue Provides a basis for exploring postmortem detection of similar heart failure-related lncRNAs miRNA for the first time used in court as evidence | Li et al.[61] |
| LINC00323 | Sensitive to hypoxia stimulation in experimental models | Potential to indicate whether hypoxia was experienced ante mortem, offering “time-of-injury inference” capability | Michalik et al.[59] |
| MIR503HG | Strongly correlated with hypoxia Remains activatable in postmortem ischemic environments | As a “hypoxia-responsive lncRNA” Can assist in determining whether myocardial ischemia existed prior to death | Michalik et al.[59] |
| Exosomal ncRNA (Nrf2/HO-1) | Exosomes derived from pericardial fluid; Elevated expression in acute ischemia | Can be used for determining cause of death in acute myocardial infarction Can be used to distinguish ante-mortem ischemia from post-mortem ischemia | Hiyamizu et al.[63] Hiyamizu et al.[63] |
Molecular basis for the forensic utility of lncRNAs
Certain lncRNAs exhibit remarkable postmortem stability in parenchymal organs, a property that significantly enhances their forensic applicability. Under hypoxic and ischemic conditions, which are frequently present in advanced HF and terminal cardiac decompensation, specific lncRNAs demonstrate sustained expression patterns that can persist after death, supporting their potential use as molecular markers in postmortem investigations.
Experimental studies have demonstrated that several intergenic lncRNAs, including LINC00323 and MIR503HG, are robustly activated in response to hypoxic stress. In the context of chronic HF, prolonged tissue hypoxia, oxidative stress, and metabolic insufficiency are hallmarks of disease progression. The hypoxia-responsive expression profiles of these lncRNAs may therefore provide time-dependent molecular information that is useful for estimating injury timing and characterizing the biological state of cardiac tissue in forensic settings[59].
Inflammation-driven cardiomyocyte injury represents a central pathological feature of end-stage HF. Transcriptomic analyses of failing hearts have revealed that differentially expressed lncRNAs are closely associated with inflammatory signaling pathways, apoptosis-related gene networks, and oxidative stress responses. The persistence of these molecular signatures after death provides a mechanistic basis for the forensic application of lncRNAs in the molecular evaluation of HF-related deaths[64].
Advantages and real-world progress of exosome-lncRNA
Emerging research has highlighted the role of EVs, including exosomes, in ncRNA-mediated cardiac processes. EVs are lipid bilayer-enclosed structures secreted by cells that mediate intercellular communication by delivering proteins, nucleic acids, and other bioactive molecules to recipient cells, thereby influencing cellular function and phenotype[65]. Exosome composition and function vary by cell type. For instance, hypoxia is a potent stimulus for exosome release from cardiomyocytes, leading to the secretion of vesicles enriched with anti-apoptotic proteins[66]. These exosomes can carry lncRNAs, such as AK139128, which modulate apoptosis and inhibit proliferation in cardiac fibroblasts, thereby contributing to repair and regeneration of infarcted myocardium[67]. Such findings highlight the dual role of exosomal lncRNAs as both biomarkers of myocardial injury and active participants in myocardial protection and remodeling, indicating their potential utility in forensic detection of heart failure.
Under conditions of hypoxia and inflammation, exosome release is further increased. This suggests that exosomal lncRNAs may help distinguish disease-induced ischemic injury from systemic ischemia occurring during the dying process, providing a potentially valuable tool for forensic diagnosis of heart failure[68]. Supporting this, Hiyamizu demonstrated that exosomes derived from pericardial fluid during autopsy contained ncRNAs, including Nrf2 and HO-1. The expression of these molecules in cardiac tissue following acute ischemia may aid postmortem diagnosis of myocardial infarction (MI)[63]. Collectively, these studies indicate that specific exosomal ncRNA markers hold significant potential for postmortem identification of heart failure.
Limitations and challenges of exosomes
Despite these advances, research on lncRNAs in forensic science remains limited relative to miRNA studies and lacks standardized methodologies. Future studies should focus on establishing uniform protocols for sample collection, exosome isolation, and RNA detection, as well as validating findings across multiple autopsy samples. Integrating lncRNA analysis with conventional cardiac biomarkers and histopathological evaluation will facilitate the development of reliable models for cardiac death determination and PMI estimation. Such efforts are expected to bridge the gap between forensic molecular diagnostics and clinical cardiovascular research, advancing both the standardization and reproducibility of postmortem RNA analysis in forensic practice.
LIMITATIONS
Despite significant advances in recent years regarding circulating lncRNA - particularly lncRNA packaged in exosomes - for the diagnosis, prognosis, and forensic applications of HF, multiple limitations remain.
First, technical protocols remain inconsistent. Differences exist across studies in exosome isolation, RNA extraction, and quantification strategies (RT-qPCR, and RNA-seq), leading to limited comparability of results. Pre-processing conditions such as temperature, centrifugation speed, and storage conditions significantly impact EV-RNA content and quality[69], while standardized detection criteria remain to be established. Second, biological origins and functional mechanisms remain unclear. Although evidence indicates that cardiomyocytes, fibroblasts, and vascular endothelial cells can release exosomes containing lncRNA, the dynamic changes and regulatory networks across different etiologies and stages of heart failure remain poorly understood[5]. Third, clinical translation and forensic applications remain in their infancy. Existing studies suffer from limited sample sizes and inconsistent sample sources - ranging from in vivo plasma samples to postmortem autopsy tissues - with no established diagnostic thresholds or reference ranges. Detection results are significantly influenced by population heterogeneity and technical conditions. lncRNAs currently cannot replace traditional protein biomarkers like BNP or NT-proBNP and serve only as supplementary indicators[13]. In forensic science, while studies indicate lncRNA detectability in early postmortem stages[70], their stability is highly influenced by tissue type, environmental temperature, and PMI. Standardized validation for exosome isolation and degradation control remains lacking[18].
FUTURE DIRECTIONS
This review aims to provide forensic scientists with a framework for establishing standardized exosome RNA extraction and quantification protocols. Combining large-scale, multi-region studies will help establish stable thresholds and reference ranges. Concurrently, multi-omics and single-cell technologies should be leveraged to uncover the dynamic regulatory networks of lncRNAs in heart failure and cardiac death. With improvements in detection sensitivity and forensic sample management standards, extracellular lncRNA holds promise as a key molecular biomarker bridging clinical heart failure diagnosis and forensic cause-of-death analysis.
In parallel, advanced multi-omics approaches, including transcriptomics, epigenomics, and proteomics, together with single-cell technologies, should be employed to elucidate the dynamic regulatory networks of lncRNAs involved in heart failure progression and cardiogenic death. Such integrative strategies may help clarify cell-type-specific lncRNA expression patterns and their mechanistic links to myocardial remodeling, fibrosis, and contractile dysfunction.
From a forensic perspective, systematic validation of postmortem lncRNA stability across different tissues, environmental conditions, and PMIs is required. The development of standardized sample handling and preservation protocols will be essential to ensure reproducibility. Ultimately, with improvements in analytical sensitivity and forensic sample management, extracellular lncRNAs may emerge as key molecular biomarkers bridging clinical heart failure diagnosis and forensic cause-of-death determination.
CONCLUSION
lncRNAs have emerged as a critical avenue for elucidating the pathophysiological mechanisms of heart failure and establishing novel molecular biomarker systems. Clinical studies indicate that circulating and exosome-encapsulated lncRNAs play crucial regulatory roles in processes such as reduced myocardial contractility and ventricular remodeling, exhibiting significant correlations with cardiac function parameters. Concurrently, forensic investigations reveal that certain lncRNAs maintain detectability and dynamic changes in postmortem samples, providing molecular evidence for cardiogenic death determination and PMI estimation.
However, current research remains constrained by inconsistent sample sources, small sample sizes, non-standardized detection protocols, and insufficient mechanistic studies. Future efforts should establish standardized exosomal RNA extraction and quantification systems, integrate transcriptomic and epigenetic data, and refine the regulatory network of heart failure-related lncRNAs. In forensic medicine, further validation of postmortem stability through multiple autopsy samples is needed to explore and establish reliable lncRNA molecular signatures, and to develop integrated models combining traditional cardiac biomarkers. With advances in high-sensitivity detection technologies and bioinformatics analysis, lncRNAs hold promise for bridging clinical and forensic medicine, offering new avenues for precise molecular identification of cardiac death.
DECLARATIONS
Authors’ contributions
Conceived the study concept and drafted the manuscript: Fan J, Li Z, Liu Q
Prepared figures and tables: Lin L, Hou J, Chai Q
Analyzed all the literature and prepared the manuscript: Wu L, Huang J, Du J
Provided supervision and reviewed the manuscript: Liu Q
Availability of data and materials
Not applicable.
AI and AI-assisted tools statement
During the preparation of this manuscript, the AI tool ChatGPT (version GPT-4o, released 2024-05-13) was used solely for language editing. The tool did not influence the study design, data collection, analysis, interpretation, or the scientific content of the work. All authors take full responsibility for the accuracy, integrity, and final content of the manuscript.
Financial support and sponsorship
This work was supported by the National Natural Science Foundation of China (grant number: 82271923).
Conflicts of interest
All authors declared that there are no conflicts of interest.
Ethical approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Copyright
© The Author(s) 2026.
REFERENCES
1. Cao Z, Jia Y, Zhu B. BNP and NT-proBNP as diagnostic biomarkers for cardiac dysfunction in both clinical and forensic medicine. Int J Mol Sci. 2019;20:1820.
2. Fan X, Zhang Z, Zheng L, Wei W, Chen Z. Long non-coding RNAs in the pathogenesis of heart failure: a literature review. Front. Cardiovasc. Med. 2022;9:950284.
4. Sato Y, Yoshihisa A, Takeishi Y. Organ injury and its management in heart failure: liver, kidney, and thyroid gland dysfunction. Fukushima J Med Sci. 2024;70:111-7.
5. Beghini A, Sammartino AM, Papp Z, et al. 2024 update in heart failure. ESC Heart Failure. 2025;12:8-42.
6. Heidenreich P. Heart failure management guidelines: new recommendations and implementation. J Cardiol. 2024;83:67-73.
7. Oremek GM, Passek K, Holzgreve F, Von Der Eltz V, Dröge J. Die biomarker BNP und NTproBNP. Zbl Arbeitsmed. 2023;73:89-95.
8. Jha S, Thasma Loganathbabu VK, Kumaran K, Krishnasamy G, Aruljothi KN. Long non-coding RNAs (lncRNAs) in heart failure: a comprehensive review. ncRNA. 2023;10:3.
9. Dhanoa JK, Sethi RS, Verma R, Arora JS, Mukhopadhyay CS. Long non-coding RNA: its evolutionary relics and biological implications in mammals: a review. J Anim Sci Technol. 2018;60:25.
10. Hinderer S, Schenke-layland K. Cardiac fibrosis - a short review of causes and therapeutic strategies. Adv Drug Delivery Rev. 2019;146:77-82.
11. Xuan L, Sun L, Zhang Y, et al. Circulating long non‐coding RNAs NRON and MHRT as novel predictive biomarkers of heart failure. J Cell Mol Med. 2017;21:1803-14.
12. Gao H, Yang S, Gao J, et al. An experimental study to estimate the early postmortem interval based on the degradation of lncRNAs in rat brain tissue. Sci Rep. 2024;14:19586.
13. Zhao J, Huang H. Extracellular vesicle-derived non-coding RNAs: key mediators in remodelling heart failure. Curr Issues Mol Biol. 2024;46:9430-48.
14. Gargiulo P, Marzano F, Salvatore M, et al. MicroRNAs: diagnostic, prognostic and therapeutic role in heart failure-a review. ESC Heart Failure. 2023;10:753-61.
15. Yan H, Ma F, Zhang Y, et al. miRNAs as biomarkers for diagnosis of heart failure: a systematic review and meta-analysis. Medicine. 2017;96:e6825.
16. Balion C, Santaguida PL, Hill S, et al. Testing for BNP and NT-proBNP in the diagnosis and prognosis of heart failure. Evid Rep Technol Assess. 2006;142:1-147.
17. Profire B, Lupașcu FG, Stătescu C, et al. Heart failure biomarkers-pathophysiology, diagnosis, prognosis and clinical relevance. Int J Mol Sci. 2025;26:9740.
18. Kim S, Jang S, Lee S, Park J, Lee S, Kim H. Characterization of exosomes and exosomal RNAs isolated from post-mortem body fluids for molecular forensic diagnosis. Diagnostics. 2022;12:2153.
19. McDonagh TA, Metra M, Adamo M, et al. 2023 focused update of the 2021 ESC guidelines for the diagnosis and treatment of acute and chronic heart failure: developed by the task force for the diagnosis and treatment of acute and chronic heart failure of the European Society of Cardiology (ESC) with the special contribution of the Heart Failure Association (HFA) of the ESC. Eur Heart J. 2023;44:3627-39.
20. Greco S, Zaccagnini G, Perfetti A, et al. Long noncoding RNA dysregulation in ischemic heart failure. J Transl Med. 2016;14:183.
21. Han P, Li W, Lin C, et al. A long noncoding RNA protects the heart from pathological hypertrophy. Nature. 2014;514:102-6.
22. Ge Z, Yin C, Li Y, et al. Long noncoding RNA NEAT1 promotes cardiac fibrosis in heart failure through increased recruitment of EZH2 to the Smad7 promoter region. J Transl Med. 2022;20:7.
23. Yu Q, Li Y, Zhang N, et al. Silencing of lncRNA NEAT1 alleviates acute myocardial infarction by suppressing miR-450-5p/ACSL4-mediated ferroptosis. Exp Cell Res. 2024;442:114217.
24. Yan L, Zhang Y, Wang M, Wang L, Zhang W, Ge Z. Circulating LIPCAR is a potential biomarker of heart failure in patients post-acute myocardial infarction. Exp Biol Med. 2021;246:2589-94.
25. Turkieh A, Beseme O, Saura O, et al. LIPCAR levels in plasma-derived extracellular vesicles is associated with left ventricle remodeling post-myocardial infarction. J Transl Med. 2024;22:31.
26. Yang Y, Mbikyo MB, Zhang J, Zhang Y, Zhang N, Li Z. The lncRNA MIAT regulates CPT-1a mediated cardiac hypertrophy through m6A RNA methylation reading protein Ythdf2. Cell Death Discov. 2022;8:167.
27. Zhang Y, Jiao L, Sun L, et al. LncRNAZFAS1 as a SERCA2a inhibitor to cause intracellular Ca2+ overload and contractile dysfunction in a mouse model of myocardial infarction. Circ Res. 2018;122:1354-68.
28. Su W, Huo Q, Wu H, et al. The function of LncRNA-H19 in cardiac hypertrophy. Cell Biosci. 2021;11:153.
29. Choong OK, Chen C, Zhang J, et al. Hypoxia-induced H19/YB-1 cascade modulates cardiac remodeling after infarction. Theranostics. 2019;9:6550-67.
30. Li J, Chen Z, Wang X, Song H. LncRNA UCA1, miR‐26a, and miR‐195 in coronary heart disease patients: correlation with stenosis degree, cholesterol levels, inflammatory cytokines, and cell adhesion molecules. J Clin Lab Anal. 2021;36:e24070.
31. Li X, Dai Y, Yan S, et al. Down-regulation of lncRNA KCNQ1OT1 protects against myocardial ischemia/reperfusion injury following acute myocardial infarction. Biochem Biophys Res Commun. 2017;491:1026-33.
32. Brunello E, Fusi L, Ghisleni A, et al. Myosin filament-based regulation of the dynamics of contraction in heart muscle. Proc. Natl. Acad. Sci. U.S.A. 2020;117:8177-86.
34. Zhang Y, Yao X-H, Wu Y, Cao G-K, Han D. LncRNA NEAT1 regulates pulmonary fibrosis through miR-9-5p and TGF-β signaling pathway. Eur Rev Med Pharmacol Sci. 2020;24:8483-92.
35. Shu L, Zhang W, Huang C, Huang G, Su G, Xu J. lncRNA ANRIL protects H9c2 cells against hypoxia‐induced injury through targeting the miR‐7‐5p/SIRT1 axis. J Cell Physiol. 2019;235:1175-83.
37. Hinton A, Claypool SM, Neikirk K, et al. Mitochondrial structure and function in human heart failure. Circ Res. 2024;135:372-96.
38. Kamradt ML, Makarewich CA. CHKB-DT: A long noncoding RNA critical for cardiovascular health. Circ Res. 2024;134:442-4.
39. Eisner DA, Caldwell JL, Kistamás K, Trafford AW. Calcium and excitation-contraction coupling in the heart. Circ Res. 2017;121:181-95.
40. Jiang D, Wang J, Wang R, Wu Y. Comprehensive insights into mechanisms for ventricular remodeling in right heart failure. Rev Cardiovasc Med. 2024;25:426.
41. Khan S, Joyce J, Margulies KB, Tsuda T. Enhanced bioactive myocardial transforming growth factor-β in advanced human heart failure. Circ J. 2014;78:2711-8.
42. Li X, Wang H, Yao B, Xu W, Chen J, Zhou X. lncRNA H19/miR-675 axis regulates cardiomyocyte apoptosis by targeting VDAC1 in diabetic cardiomyopathy. Sci Rep. 2016;6:36340.
43. Wang Z, Zhang X, Ji Y, et al. The long noncoding RNA Chaer defines an epigenetic checkpoint in cardiac hypertrophy. Nat Med. 2016;22:1131-9.
44. Viereck J, Kumarswamy R, Foinquinos A, et al. Long noncoding RNA Chast promotes cardiac remodeling. Sci Transl Med. 2016;8.
45. Van Heesch S, Witte F, Schneider-lunitz V, et al. The translational landscape of the human heart. Cell. 2019;178:242-260.e29.
46. Yan Y, Tang R, Li B, et al. The cardiac translational landscape reveals that micropeptides are new players involved in cardiomyocyte hypertrophy. Mol Ther. 2021;29:2253-67.
47. Barbalata T, Niculescu LS, Stancu CS, Pinet F, Sima AV. Elevated levels of circulating lncRNAs LIPCAR and MALAT1 predict an unfavorable outcome in acute coronary syndrome patients. Int J Mol Sci. 2023;24:12076.
48. Qi X, Yu L, Liu S, et al. The expression profile analysis and functional prediction of lncRNAs in peripheral blood mononuclear cells in maintenance hemodialysis patients developing heart failure. Sci Rep. 2024;14:29577.
49. Ma Y, Brocchini S, Williams GR. Extracellular vesicle-embedded materials. J Control Release. 2023;361:280-96.
50. Willms E, Johansson HJ, Mäger I, et al. Cells release subpopulations of exosomes with distinct molecular and biological properties. Sci Rep. 2016;6:22519.
51. Berumen Sánchez G, Bunn KE, Pua HH, Rafat M. Extracellular vesicles: mediators of intercellular communication in tissue injury and disease. Cell Commun Signal. 2021;19:104.
52. Gezer U, Özgür E, Cetinkaya M, Isin M, Dalay N. Long non‐coding RNAs with low expression levels in cells are enriched in secreted exosomes. Cell Biol Int. 2014;38:1076-9.
53. Kim KM, Abdelmohsen K, Mustapic M, Kapogiannis D, Gorospe M. RNA in extracellular vesicles. WIREs RNA. 2017;8:e1413.
54. Wang L, Zhang Y, Wang J, et al. Exosomal lncRNA profiles in patients with HFrEF: evidence for KLF3-AS1 as a novel diagnostic biomarker. Mol Cell Probes. 2025;82:102032.
55. Li J, Wu X, Ma H, et al. New developments in non-exosomal and exosomal ncRNAs in coronary artery disease. Epigenomics. 2022;14:1355-72.
56. Zhao L, Gu M, Sun Z, et al. The role of exosomal lncRNAs in cardiovascular disease: emerging insights based on molecular mechanisms and therapeutic target level. Non-coding RNA Res. 2025;10:198-205.
57. Liu Y, Wang C, Fu X, Ren M. The progress and evolving trends in nucleic-acid-based therapies. Biomolecules. 2025;15:376.
58. Kim Y. RNA therapy: rich history, various applications and unlimited future prospects. Exp Mol Med. 2022;54:455-65.
59. Michalik KM, You X, Manavski Y, et al. Long noncoding RNA MALAT1 regulates endothelial cell function and vessel growth. Circ Res. 2014;114:1389-97.
60. Deiman FE, De Graaf MM, Sillje HHW, Grote Beverborg N, Bomer N, Van Der Meer P. RNA therapeutics in heart failure. J Cardiovasc Transl Res. 2025;18:1540-54.
61. Li Y, Wang Z, Ishmael D, Lvy Y. The potential of using non-coding RNAs in forensic science applications. Forensic Sci Res. 2023;8:98-106.
62. Hanson EK, Lubenow H, Ballantyne J. Identification of forensically relevant body fluids using a panel of differentially expressed microRNAs. Anal Biochem. 2009;387:303-14.
63. Hiyamizu S, Ishida Y, Yasuda H, et al. Forensic significance of intracardiac expressions of Nrf2 in acute myocardial ischemia. Sci Rep. 2024;14:4046.
64. Frangogiannis NG. The extracellular matrix in myocardial injury, repair, and remodeling. J Clin Investig. 2017;127:1600-12.
65. Tkach M, Théry C. Communication by extracellular vesicles: where we are and where we need to go. Cell. 2016;164:1226-32.
66. Chistiakov D, Orekhov A, Bobryshev Y. Cardiac extracellular vesicles in normal and infarcted heart. Int J Mol Sci. 2016;17:63.
67. Wang L, Zhang J. Exosomal lncRNA AK139128 derived from hypoxic cardiomyocytes promotes apoptosis and inhibits cell proliferation in cardiac fibroblasts. Int J Nanomedicine. 2020;15:3363-76.
68. Bellin G, Gardin C, Ferroni L, et al. Exosome in cardiovascular diseases: a complex world full of hope. Cells. 2019;8:166.
69. Bernáth-nagy D, Kalinyaprak MS, Giannitsis E, et al. Circulating extracellular vesicles as biomarkers in the diagnosis, prognosis and therapy of cardiovascular diseases. Front. Cardiovasc. Med. 2024;11:1425159.
Cite This Article
How to Cite
Download Citation
Export Citation File:
Type of Import
Tips on Downloading Citation
Citation Manager File Format
Type of Import
Direct Import: When the Direct Import option is selected (the default state), a dialogue box will give you the option to Save or Open the downloaded citation data. Choosing Open will either launch your citation manager or give you a choice of applications with which to use the metadata. The Save option saves the file locally for later use.
Indirect Import: When the Indirect Import option is selected, the metadata is displayed and may be copied and pasted as needed.
About This Article
Special Topic
Copyright

Data & Comments
Data
0











Comments
Comments must be written in English. Spam, offensive content, impersonation, and private information will not be permitted. If any comment is reported and identified as inappropriate content by OAE staff, the comment will be removed without notice. If you have any queries or need any help, please contact us at support@oaepublish.com.