




Stem cell extracellular vesicles for neuropsychiatric disorders and translation
 0
0 Abstract
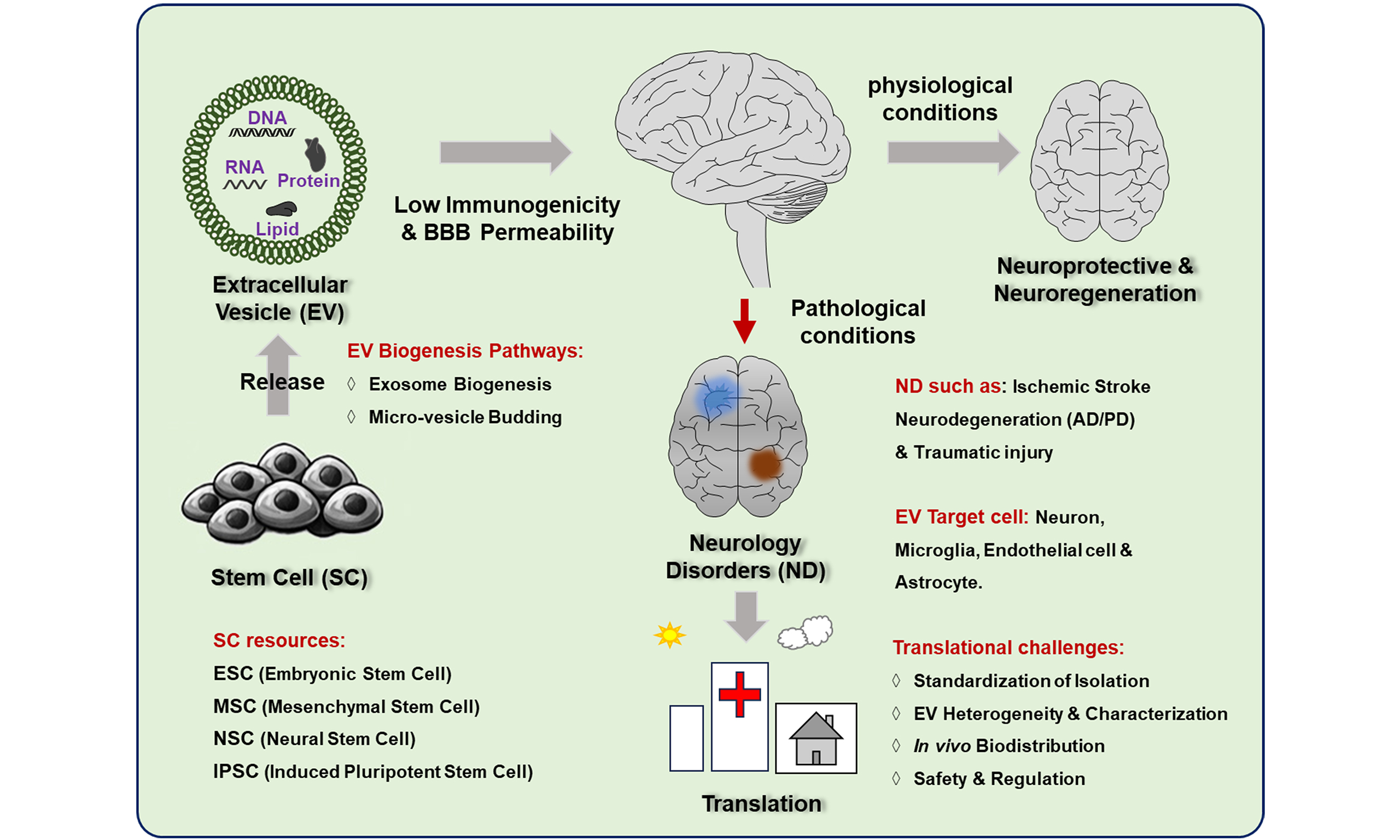
Neuropsychiatric disorders represent a major global health challenge due to their high prevalence, chronic disability, and substantial socioeconomic burden. Although stem cell-based therapies offer regenerative potential, their clinical application is limited by poor post-transplantation survival, restricted targeted integration, and potential tumorigenicity. Stem cell-derived extracellular vesicles (SC-EVs), particularly exosomes, have emerged as a promising cell-free therapeutic approach. These vesicles can cross the blood-brain barrier (BBB) and exhibit high biocompatibility and low immunogenicity. This review summarizes the cellular origins and biogenesis of SC-EVs and evaluates current preclinical and clinical evidence supporting their therapeutic potential. Particular attention is given to acute ischemic stroke and progressive neurodegenerative disorders, including Alzheimer’s disease and Parkinson’s disease. In addition, the molecular mechanisms underlying their neuroprotective and regenerative effects are discussed, with a focus on modulating neuroinflammation, promoting neurogenesis, and enhancing synaptic plasticity. Finally, key advances and major challenges in the clinical translation of SC-EVs are outlined. Integrating current evidence, this review provides a framework and practical perspective for the continued development of SC-EV-based therapies for complex neurological disorders.
Keywords
INTRODUCTION
Neurological disorders are a major global public health challenge, affecting approximately 3.4 billion people and causing nearly 11.8 million deaths annually, thereby imposing a heavy socioeconomic burden[1]. As the global population ages, the incidence of these disorders continues to rise, placing increasing pressure on healthcare systems[2]. A major obstacle to effective neurotherapeutics is the blood-brain barrier (BBB), which prevents most small molecules and nearly all macromolecular biologics from entering the central nervous system (CNS)[3]. In addition, neuronal damage in these disorders is complex, heterogeneous, and often irreversible, which limits the effectiveness of conventional monotherapies[4]. These challenges highlight the need for novel, multifaceted regenerative strategies.
Stem cells, characterized by self-renewal, proliferation, and multilineage differentiation, are considered promising candidates for the treatment of neurological disorders[5,6]. Common types include mesenchymal stem cells (MSCs), neural stem cells (NSCs), induced pluripotent stem cells (iPSCs), and embryonic stem cells (ESCs). These cells exert therapeutic effects through multiple mechanisms, including cell replacement, immunomodulation[7], anti-inflammatory effects, promotion of angiogenesis, and enhancement of neuroprotection[8]. However, increasing evidence indicates that their therapeutic benefits are largely mediated by paracrine signaling rather than direct in vivo engraftment[9].
Despite their therapeutic potential, current stem cell-based therapies face translational challenges. Intrinsic cellular heterogeneity complicates controlled differentiation and the standardization of clinical outcomes[10]. Moreover, poor post-transplantation survival and poor homing to injury sites limit efficacy, and there is a risk of immune rejection in allogeneic settings[11]. In addition, undifferentiated stem cells may form tumors, so careful safety measures such as rigorous purification and phenotypic validation are required before clinical use[12].
In this context, stem cell-derived extracellular vesicles (SC-EVs) have emerged as a promising cell-free alternative. SC-EVs retain key regenerative and immunomodulatory components of their parent cells while avoiding many of the safety, immunological, and engraftment limitations associated with live-cell transplantation[5,6]. Although previous studies have summarized general aspects of extracellular vesicle biology, this review provides a focused and clinically oriented overview of SC-EV-mediated neurological repair, organized into six principal mechanistic categories: immunomodulation, mitochondrial homeostasis, neuroprotection, enhancement of neuroplasticity, angiogenesis, and clearance of pathological protein aggregates.
This review focuses primarily on conditions supported by strong preclinical and translational evidence, including acute ischemic injury (stroke) and progressive neurodegenerative disorders such as Alzheimer’s disease and Parkinson’s disease (PD). Emerging applications in neuropsychiatric disorders (e.g., epilepsy, schizophrenia, and major depressive disorder) are also discussed. These conditions are increasingly recognized not only as disorders of neurotransmission but also as involving structural and neuroinflammatory alterations in the CNS, making them relevant targets for SC-EV-based interventions. Finally, to bridge the gap between experimental research and clinical application, we evaluate the current clinical trial landscape and highlight key methodological challenges, including EV heterogeneity, isolation-related biases, and the lack of standardized characterization criteria that must be addressed to enable reliable clinical translation.
LITERATURE SEARCH STRATEGY
A comprehensive literature search [Figure 1] was conducted to identify relevant studies on SC-EVs in neurological disorders. The major electronic databases, including PubMed, Web of Science, and Scopus, were systematically searched for articles published from 2015 to 2025. The search strategy combined keyword and Medical Subject Headings (MeSH) terms related to “stem cell-derived extracellular vesicles,” “isolation and purification,” and specific neurological disorders (e.g., stroke, epilepsy, schizophrenia, depression, neuropsychiatric diseases, neurodegenerative diseases).
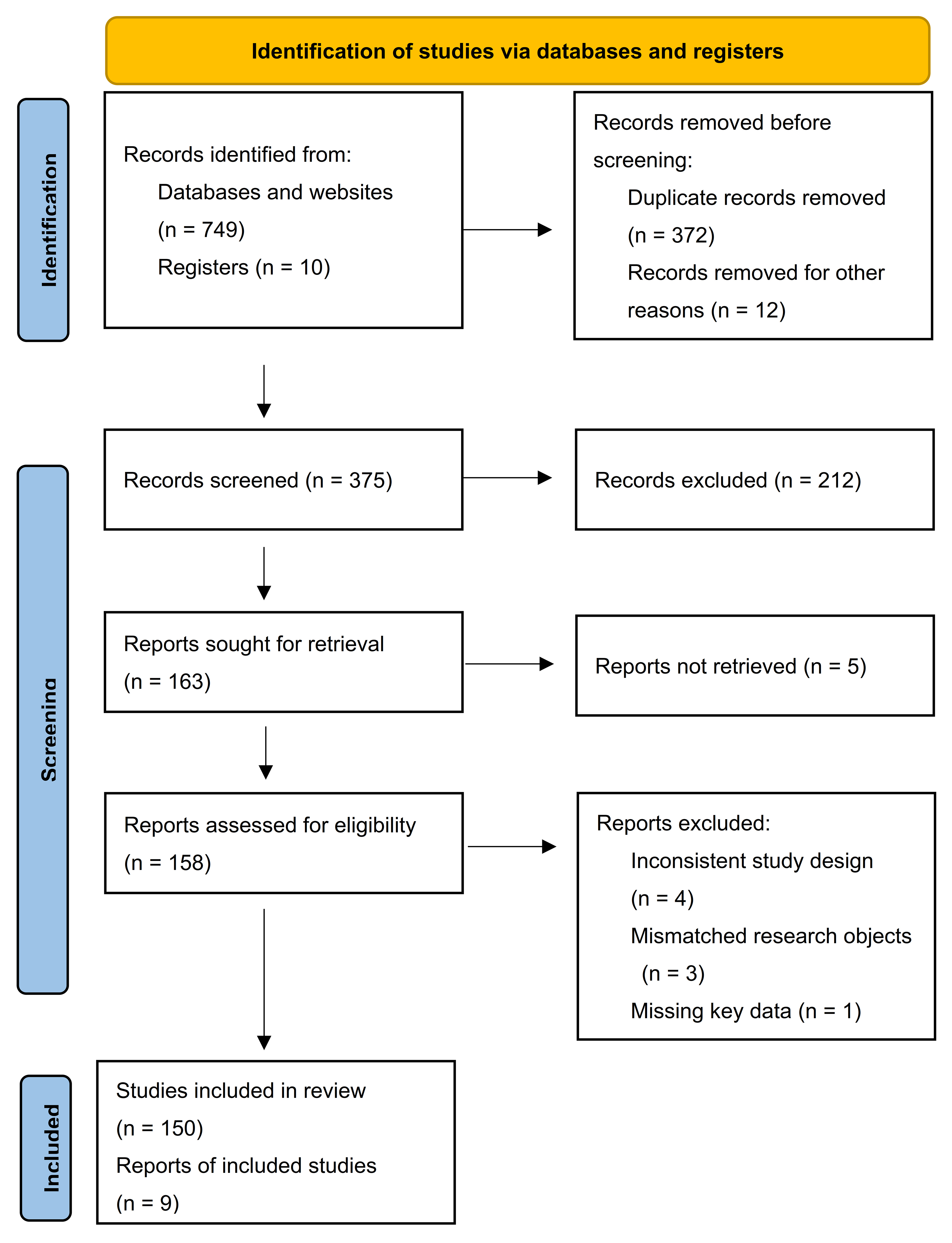
Figure 1. PRISMA flow diagram. This study followed the PRISMA 2020 guidelines for systematic literature screening. The initial search of databases and clinical trial registries yielded 759 records (749 from databases and websites and 10 from registries). After removing 372 duplicate records and 12 records excluded for other reasons, 375 records remained for screening. Title and abstract screening excluded 212 records that did not meet the inclusion criteria. Of the 163 records selected for full-text review, 5 could not be retrieved, leaving 158 articles for eligibility assessment. At this stage, 8 articles were excluded due to inconsistent study design, inappropriate study populations, or missing essential data. Ultimately, 150 records (148 journal articles and 2 registry entries) were included in final evidence synthesis. PRISMA: Preferred Reporting Items for Systematic reviews and Meta-Analyses.
Eligible studies included original research articles, preclinical studies, and clinical investigations that evaluated the therapeutic potential or underlying mechanisms of SC-EVs in neurological contexts. Review articles were also screened to identify additional relevant references. Studies not published in English or lacking sufficient methodological detail were excluded.
Two independent reviewers screened titles and abstracts for relevance, followed by full-text assessment of eligible articles. Discrepancies were resolved through discussion and consensus. Data extraction focused on study design, source of stem cells, EV isolation methods, experimental models, therapeutic outcomes, and mechanistic findings.
SC-EVS: BIOGENESIS, SECRETION AND CELLULAR SOURCE
Overview of SC-EVs
SC-EVs are nanoscale, membrane-bound vesicles (typically 30-150 nm in diameter) released by cells and involved in intercellular communication[13]. Their biological activity is mediated by cargo that includes proteins, lipids, and nucleic acids[14]. Proteomic analyses have identified enrichment of specific surface receptors, cytoskeletal components, and signaling molecules within these vesicles[15].
According to the Minimal Information for Studies of Extracellular Vesicles (MISEV) guidelines[16,17], the characterization of SC-EVs requires a multiparametric approach. Morphology and size distribution should be assessed using complementary high-resolution techniques, such as transmission electron microscopy (for morphology) and nanoparticle tracking analysis or tunable resistive pulse sensing (for size distribution and concentration). Quantification should include both total particle number and total protein content, as the particle-to-protein ratio provides an indirect measure of sample purity.
To confirm vesicle identity, protein profiling should demonstrate the presence of at least one transmembrane or lipid-associated marker (e.g., CD9, CD63, CD81) together with a cytosolic EV-associated protein [e.g., tumor susceptibility gene 101 (TSG101) or ALG-2-interacting protein X (Alix)]. In addition, at least one negative marker (e.g., calnexin for the endoplasmic reticulum or GM130 for the Golgi apparatus) should be evaluated to exclude contamination from intracellular organelles. Screening for common non-vesicular contaminants, including lipoproteins and protein aggregates, is also necessary to ensure that observed functional effects are attributable to EVs. Because no single method can fully define EV identity or purity, the combined use of multiple approaches is recommended[17].
In addition to proteins, SC-EVs contain nucleic acids, including messenger ribonucleic acid (mRNA), micro ribonucleic acid (miRNA), and DNA fragments. Among these components, miRNAs are particularly important because they regulate gene expression and mediate intercellular communication. The miRNA composition of SC-EVs varies according to the cellular source, resulting in distinct regulatory profiles[17,18]. The lipid composition of SC-EVs also contributes to their biological activity. Enrichment of cholesterol, sphingolipids, and phosphatidylserine supports membrane stability and facilitates processes such as membrane fusion and endocytosis. Lipidomic studies further indicate that source-specific lipid profiles influence cargo loading and target cell interactions[19-22].
Biogenesis and release of SC-EVs
SC-EVs are formed through the maturation of multivesicular bodies (MVBs). During this process, the endosomal membrane invaginates to encapsulate selected cytosolic components, forming intraluminal vesicles (ILVs). These MVBs subsequently fuse with the plasma membrane, releasing ILVs into the extracellular space as vesicles enriched in tetraspanins such as CD63[23].
The sorting and budding of vesicles are primarily regulated by the endosomal sorting complex required for transport (ESCRT) machinery. This system consists of four sub-complexes (ESCRT-0, -I, -II, and -III), which function sequentially to recognize ubiquitinated cargo, induce membrane invagination, and mediate vesicle scission. Accessory proteins such as hepatocyte growth factor-regulated tyrosine kinase substrate (HRS) and TSG101 facilitate cargo clustering, while VPS4 mediates membrane scission and complex disassembly[24].
In addition to ESCRT-dependent pathways, SC-EVs can also be generated through ESCRT-independent mechanisms. One major pathway involves ceramide generation by neutral sphingomyelinases, which induces membrane curvature and promotes ILV formation. This process can be inhibited by neutral sphingomyelinase inhibitors such as GW4869. Tetraspanins (such as CD9, CD63, CD81) also contribute to cargo organization through membrane microdomains, independent of ubiquitination. Other proteins, including SIMPLE and Ndfip1, participate in cargo recruitment[23]. The coexistence of multiple biogenesis pathways within a single cell contributes to the heterogeneity of EV subpopulations [Figure 2].
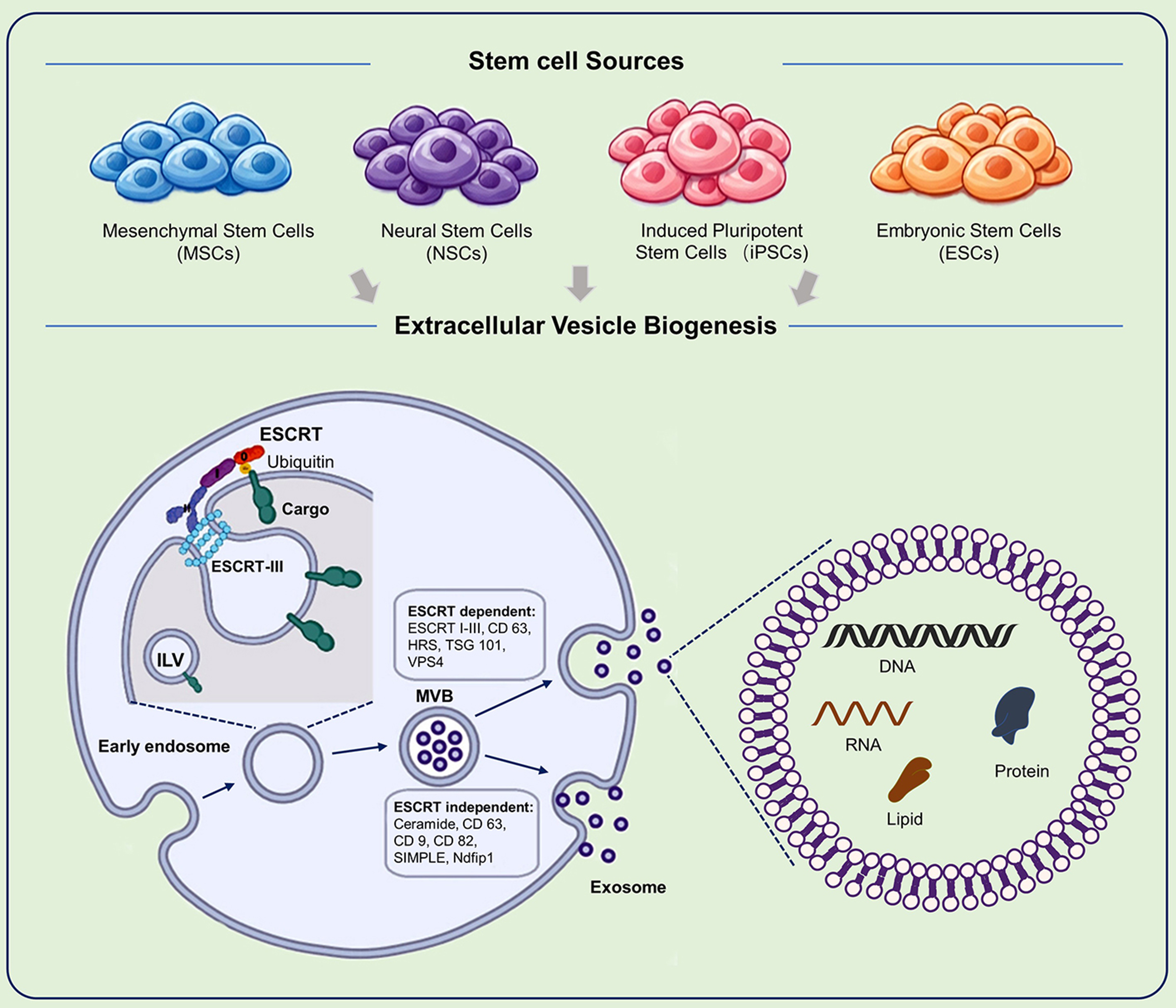
Figure 2. Biogenesis and Release of SC-EVs. This schematic illustrates the cellular pathways involved in the formation, maturation, and secretion of SC-EVs. The upper panel represents the parent stem cell, whereas the lower panel depicts key stages of vesicle biogenesis. The process begins with invagination of the plasma membrane to form early endosomes, which subsequently mature into late endosomes or MVBs. During this process, inward budding of the endosomal membrane encapsulates selected proteins, lipids, and nucleic acids within ILVs. Cargo sorting is regulated by both ESCRT-dependent and ESCRT-independent mechanisms. Upon maturation, MVBs fuse with the plasma membrane, releasing ILVs into the extracellular space as exosomes. In contrast, microvesicles are generated through direct outward budding and fission of the plasma membrane. SC-EVs: Stem cell-derived extracellular vesicles; ESCRT: endosomal sorting complex required for transport; HRS: hepatocyte growth factor-regulated tyrosine kinase substrate; TSG101: tumor susceptibility gene 101; VPS4: vacuolar protein sorting-associated protein 4; ILV: intraluminal vesicle; MVB: multivesicular body; SIMPLE:small integral membrane protein of the endosome; Ndfip1: Nedd4 family interacting protein 1; DNA: deoxyribonucleic acid; RNA: ribonucleic acid.
Cellular sources of SC-EVs
MSCs
MSCs are among the most extensively studied sources of therapeutic extracellular vesicles. Although MSC-derived EVs express canonical tetraspanins (CD9, CD63, and CD81), their secretomes exhibit substantial heterogeneity[25-29], which affects their functional properties, as does methodological variation during isolation[30-32].
Among MSC subtypes, bone marrow-derived MSCs (BMSCs) show strong neuroprotective potential but are limited by invasive harvesting procedures and batch variability[15,33-37]. In contrast, umbilical cord-derived MSCs (UCMSCs) provide a scalable and low-immunogenic alternative that is suitable for clinical applications[36,38-42]. Adipose-derived MSCs (ADMSCs) offer high yield through minimally invasive collection[43-46]; however, their isolation requires careful enzymatic processing to avoid altering EV composition[15,47,48].
Pluripotent stem cells
Embryonic stem cell-derived EVs (ESC-EVs) contain potent regenerative cargo; however, their clinical use is limited by ethical concerns and the risk of teratoma formation[49-55]. iPSCs provide a more practical alternative for personalized therapy[37,56]. iPSC-derived EVs are enriched in developmental miRNAs and regulatory factors that promote angiogenesis and tissue repair. However, because their secretome can be highly dynamic, careful isolation and characterization are required to ensure consistency and reproducibility.
Neural stem and progenitor cells
Neural stem/progenitor cells (NSCs/NPCs) produce EVs that are suited for CNS repair[57]. These vesicles are enriched in neural-specific miRNAs and synapse-associated proteins[58-60], supporting neuroprotection, anti-inflammatory effects, and axonal regeneration[61,62]. Large-scale production of NSC-derived EVs often relies on high-throughput methods such as Tangential flow filtration (TFF) for EV concentration; careful validation is required to preserve EV integrity and function.
Isolation of SC-EVs
The isolation of SC-EVs from conditioned media remains a key technical challenge. Consistent with MISEV guidelines, no single method can be considered universally optimal; instead, each approach introduces specific biases that influence the composition of recovered EV populations[63-65]. Differential ultracentrifugation is widely used but may cause vesicle aggregation and co-isolation of contaminants such as protein complexes and lipoproteins. Size-exclusion chromatography (SEC) improves purity and preserves vesicle integrity; however, it often results in sample dilution and may require additional concentration steps[64-66]. Density gradient centrifugation, particularly with iodixanol, enables high-resolution separation based on buoyant density and reduces contamination from other similarly sized particles[64,67].
Immunoaffinity capture using antibodies against surface markers (e.g., CD9 or CD63) allows selective enrichment of EV subpopulations but is limited by low throughput and potential interference with surface proteins[64,68]. Overall, the choice of isolation method influences EV composition and functional outcomes, thus highlighting the need for standardized characterization [Table 1].
Methodological overview for stem cell-derived extracellular vesicle isolation
| Method/Technique | Principle | Key advantages | Major limitations |
| Differential ultracentrifugation | Sequential centrifugation based on size/density sedimentation | Widely applicable[69,70] | Time-consuming; requires specialized equipment; Non-specific co-precipitation; vesicle damage/aggregation[71-73] |
| Size-exclusion chromatography | Separation by hydrodynamic volume using a porous column matrix | High purity; preserves vesicle integrity and function[74,75] | Moderate throughput; sample dilution[17,76,77] |
| Ultrafiltration | Size-based separation using membranes with defined molecular weight cut-offs | Scalable; suitable for processing large volumes[78] | Risk of vesicle deformation or membrane clogging[79] |
| Density gradient centrifugation | Separation based on buoyant density within a gradient medium | High-resolution separation from contaminants of similar sizes[64,67,80] | Labor-intensive; requires long centrifugation times[81] |
| Immunoaffinity capture | Selective binding using antibodies targeting specific surface markers | High specificity for distinct subpopulations or sources[82] | High cost; potential for antibody contamination; low throughput[64,68,79] |
PRECLINICAL RESEARCH OF EVS IN NEUROLOGICAL DISORDERS
Immunomodulation and anti-neuroinflammation
The modulation of neuroinflammation is a core mechanism underlying the therapeutic effects of SC-EVs across neurological disorders. These effects involve regulation of glial activation states, suppression of pro-inflammatory mediators, and promotion of immune phenotypes associated with tissue repair.
Stroke: modulation of microglial dynamics and inflammatory cascades
In ischemic stroke, SC-EVs deliver molecular cargo that modulates inflammatory cascades and mitigates ischemia-reperfusion (I/R) injury. A key mechanism involves promoting a shift in microglia from a pro-inflammatory (M1-like) state toward a neuroprotective (M2-like) phenotype. For example, BMSC-EVs enriched in the lncRNA zinc finger antisense 1 (ZFAS1) reduce microglial activation by downregulating miR-15a-5p[83], while other EV populations suppress the NLR family pyrin domain containing 3 (NLRP3) inflammasome to attenuate neuronal pyroptosis[84]. SC-EVs also influence post-transcriptional and post-translational regulation. BMSC-EVs carrying the lncRNA Krüppel-like factor 3 antisense RNA 1 (KLF3-AS1) inhibit sirtuin 1 (Sirt1) ubiquitination via USP22, alleviating I/R-induced injury[85]. In addition, UCMSC-EVs modulate microglial activity through high-mobility group box 1 (HMGB1)-mediated activation of the triggering receptor expressed on myeloid cells 1 (TREM1)-p38 mitogen-activated protein kinase (MAPK) pathway, whereas NSC-EVs deliver miR-125a-5p to inhibit the toll-like receptor 4 (TLR4)/nuclear factor-kappa B (NF-κB) signaling axis[86-88]. SC-EVs also reduce oxidative stress, for example by targeting the fat mass and obesity-associated protein (FTO)/N6-methyladenosine (m6A)/ dynamin-related protein (Drp1) pathway, which indirectly attenuates inflammation-associated neuronal injury[89].
Neurodegenerative diseases: restoration of immune homeostasis and microglial balance
In Alzheimer’s disease (AD), SC-EVs reduce amyloid-β (Aβ) accumulation and associated glial activation, leading to decreased expression of pro-inflammatory cytokines such as tumor necrosis factor-alpha (TNF-α), interleukin-1 beta (IL-1β), and interleukin-6 (IL-6)[90]. In parallel, they increase brain-derived neurotrophic factor (BDNF) expression[91] and promote a shift in microglial phenotype toward a reparative state. These changes may partially counteract age-associated inflammatory processes[92,93] and reduce reactive astrogliosis[94].
In PD, SC-EVs suppress inflammatory signaling in the substantia nigra, which is important for protecting dopaminergic neurons. Engineered BMSC-EVs (e.g., AK76 or quercetin-loaded EVs) promote M2-like polarization[95], while Gli1-containing EVs inhibit Sp1-mediated activation of Leucine-rich repeat kinase 2 (LRRK2)[96]. UCMSC-EVs are preferentially internalized by activated microglia in lesion areas, contributing to localized immunomodulation and preservation of neuronal integrity[97,98].
Emerging applications in neuropsychiatric disorders
In epilepsy, SC-EVs affect the interaction between neuroinflammation and neuronal hyperexcitability. ADMSC-EVs deliver miR-23b-3p to regulate the signal transducer and activator of transcription 1 (STAT1)/Glyoxylate reductase 1 homolog (GLYR1) axis and promote microglial phenotype transition[99,100], while UCMSC-EVs activate nuclear factor erythroid 2-related factor 2 (NRF2) signaling to inhibit NLRP3 inflammasome activation[101,102]. iPSC-EVs also suppress inflammatory signaling via the lncRNA-0949-TLR4/MAPK/NF-κB pathway[103].
In psychiatric disorder models, SC-EVs modulate inflammatory balance. In schizophrenia, MSC-EVs reduce pro-inflammatory cytokine expression while increasing IL-10 levels[104]. In depression models, UCMSC-EVs inhibit the myeloid differentiation primary response 88 (MyD88)/tumor necrosis factor receptor-associated factor 6 (TRAF6)/NF-κB pathway via miR-125b-5p, thereby reducing neuroinflammation and synaptic impairment[105]. ADMSC-EVs also activate adenosine monophosphate-activated protein kinase (AMPK)-dependent autophagy and suppress NLRP3 signaling[106].
Critical appraisal and translational gaps
Although SC-EVs consistently attenuate neuroinflammation in preclinical studies, several limitations affect translational interpretation. First, many studies rely on the simplified M1/M2 polarization framework, whereas microglial activation in vivo represents a continuous and heterogeneous spectrum, including disease-associated microglia (DAM). Second, experimental models often focus on microglia in isolation, with limited consideration of interactions with astrocytes, endothelial cells, and peripheral immune components. Finally, variability in cytokine measurement and experimental design reduces cross-study comparability and reproducibility.
Regulation of oxidative stress and mitochondrial homeostasis
Oxidative stress and mitochondrial dysfunction are common features of ischemic, neurodegenerative, and psychiatric disorders. SC-EVs reduce lipid peroxidation and support mitochondrial function through delivery of regulatory RNAs and proteins.
Stroke and PD: targeted modulation of redox status and mitophagy
In ischemic stroke, SC-EVs stabilize mitochondrial dynamics. For example, BMSC-EVs regulate the Krüppel-like factor 4 (KLF4)-FTO pathway and reduce the m6A modification of Drp1, thereby limiting mitochondrial fragmentation[89]. SC-EVs also reduce reactive oxygen species (ROS) production through mechanisms involving the lncRNA ZFAS1[83] and phosphatidylinositol 3-kinase (PI3K)/protein kinase B (AKT)/mammalian target of rapamycin (mTOR) signaling[88]. In addition, NSC-EVs activate PINK1/Parkin-mediated mitophagy, promoting removal of damaged mitochondria[107].
Similar mechanisms are observed in PD models. SC-EVs reduce oxidative stress in the substantia nigra and protect dopaminergic neurons. Both NSC-EVs[98] and engineered BMSC-EVs[95] decrease ROS levels and reduce neuronal loss in toxin-induced models.
Depression: antioxidant signaling and neuroprotection
Depression is increasingly associated with chronic oxidative stress and metabolic dysfunction alongside classical neurotransmitter imbalance. In this context, the therapeutic effects of SC-EVs are partly mediated by regulating redox homeostasis. BMSC-EVs increase hippocampal miR-26a levels and reduce oxidative damage markers, including malondialdehyde and nitric oxide[108]. In parallel, SC-EVs modulate downstream signaling pathways such as MAPK and NF-κB, thereby reducing apoptosis and supporting synaptic function[109]. These findings suggest that regulation of mitochondrial function and oxidative stress contributes to the neuroprotective effects of SC-EVs in depression models.
Critical appraisal and translational gaps
Despite consistent antioxidant effects in preclinical studies, several limitations remain. Most experimental models rely on acute oxidative stress paradigms that fail to reflect the chronic and progressive nature of mitochondrial dysfunction in human disease. In addition, the transient nature of ROS, together with variability in detection methods, complicates cross-study comparisons. The limited availability of in vivo tracking approaches makes it difficult to determine whether administered SC-EVs effectively reach target cells and interact with mitochondrial pathways.
Neuronal protection and anti-apoptotic signaling
Programmed cell death is a common feature of both acute brain injury and chronic neurodegeneration. SC-EVs exert neuroprotective effects by modulating apoptotic pathways, reducing excitotoxicity, and preserving neuronal structure and function.
Stroke and AD: suppression of cell death pathways
In ischemic stroke, SC-EVs influence multiple forms of cell death. ADMSC-EVs deliver miR-25-3p to hypoxic neurons, suppressing p53 and the B-cell lymphoma 2 (BCL2)-interacting protein 3, thereby restoring autophagic balance and reducing apoptosis[110]. They also inhibit ferroptosis by targeting CHAC1 via miR-760-3p[111]. In addition, UCMSC-EVs regulate PI3K/AKT and MAPK signaling, decreasing pro-apoptotic factors (Bax, Caspase-3) while increasing anti-apoptotic Bcl-2 expression[112].
In AD models, SC-EVs reduce amyloid-associated neurotoxicity and support neuronal survival. BMSC-EVs increase expression of neuronal and neurogenic markers [neuronal nuclei (NeuN), NeuroD1, BDNF][91,113,114], while UCMSC-EVs reduce Aβ burden and partially restore synaptic gene expression[115]. NSC-EVs also activate Sirt1 signaling to support mitochondrial function under amyloid stress[94].
PD, epilepsy, and schizophrenia: targeted neuroprotection
In PD models, SC-EVs protect dopaminergic neurons through metabolic and autophagy-related mechanisms. Engineered BMSC-EVs (AK76 and quercetin) promote autophagy[95], while EV-associated tumor necrosis factor stimulated gene 6 protein (TSG 6) reduces 1-methyl-4-phenylpyridinium toxicity via the STAT3/miR-7/neural precursor cell expressed developmentally down regulated protein 4 (NEDD4) pathway[116]. In epilepsy, EV-delivered circRNA hsa_circ_0000288 regulates cell cycle associated protein 1 (CAPRIN1)/glutamate receptor ionotropic NMDA subunit 3B (NR3B) signaling, reducing hippocampal damage[117]. ADMSC-EVs also inhibit NLRP3-mediated pyroptosis[118]. In schizophrenia models, MSC-EVs protect parvalbumin-positive interneurons and help maintain inhibitory circuit stability[119].
Critical appraisal and translational gaps
Although SC-EVs reduce markers of apoptosis in preclinical studies, important limitations remain. Most models are based on acute injury paradigms that do not reflect the gradual and region-specific neuronal loss observed in human disease. In addition, many studies rely on short-term or static apoptosis markers, which may not predict long-term neuronal survival or functional integration. Therefore, it remains unclear whether these short-term protective effects translate into sustained functional recovery.
Regulation of neurogenesis and synaptic plasticity
Impaired neurogenesis and synaptic dysfunction contribute to the progression of multiple neurological disorders. SC-EVs have been reported to promote neural progenitor proliferation, synaptic repair, and plasticity.
Schizophrenia and depression: neurogenesis and synaptic repair
In schizophrenia, MSC-EVs increase the number of doublecortin-positive cells in the hippocampus, suggesting reactivation of neurogenic pathways, potentially through restoration of BDNF signaling. They also upregulate the postsynaptic density protein 95 (PSD-95) and tyrosine hydroxylase, contributing to the stabilization of synaptic structure and dopaminergic neurotransmission[104]. Similar effects have been reported in depressive disorders. BMSC-EVs activate the BDNF/GDNF signaling axis and promote neuronal proliferation in the dentate gyrus (DG)[120]. NSC-EVs, particularly when combined with electroacupuncture, enhance synaptic protein expression, including PSD-95, synaptophysin (SYN), and growth-associated protein 43, thereby facilitating synaptic remodeling[121]. In addition, ADMSC-EVs inhibit Galectin-3-mediated microglial synaptic pruning, which may help preserve synaptic connectivity in the prefrontal cortex[122].
AD and stroke: restoration of plasticity and network connectivity
In both neurodegenerative and ischemic conditions, SC-EVs contribute to the restoration of neural network integrity. In AD, NSC-EVs and UCMSC-EVs have been shown to partially normalize the expression of genes involved in synaptic plasticity[94,115], which correlates with improved cognitive performance in amyloid precursor protein (APP)/presenilin 1 (PS1) models.
In stroke, SC-EVs support both neurogenesis and structural remodeling in peri-infarct regions. ADMSC-EVs engineered with PD-L1 and hepatocyte growth factor activate the STAT3/forkhead box O3 (FOXO3) pathway and promote neurogenesis[123]. In parallel, NSC-EVs enhance axonal structure and facilitate synaptic reconnection in the ischemic penumbra, contributing to functional recovery[88,107]. These effects are particularly relevant given that post-stroke recovery largely depends on the plasticity of surviving neural tissue.
Critical appraisal and translational gaps
Although SC-EVs promote neurogenesis and synaptic remodeling in preclinical models, significant translational challenges remain. Species differences in adult neurogenesis limit direct extrapolation to humans. In addition, it remains unclear whether EV-induced neurons and newly formed synapses can achieve stable, long-term functional integration within existing neural circuits in vivo.
Angiogenesis and BBB integrity
Vascular dysfunction and BBB disruption are key features of both acute ischemic injury and chronic neurodegeneration. Rather than acting solely on neurons, SC-EVs also influence the neurovascular unit by promoting angiogenesis and stabilizing BBB structure.
Stroke: revascularization and barrier defense
In ischemic stroke, therapeutic strategies must balance reperfusion with the risk of secondary injury caused by BBB disruption. SC-EVs contribute to both processes. BMSC-EVs, particularly under hypoxic preconditioning, carry pro-angiogenic microRNAs such as miR-126-3p and miR-140-5p, which enhance endothelial cell migration and tube formation[124]. This response is further supported by increased expression of angiogenic factors, including vascular endothelial growth factor (VEGF), vascular endothelial growth factor receptor 2 (VEGFR2), angiopoietin-1 (Ang-1), tyrosine kinase with immunoglobulin and epidermal growth factor homology domains 2 (Tie-2)[125].
In addition, SC-EVs help maintain BBB integrity. BMSC-EVs inhibit Caveolin-1-mediated internalization of tight junction proteins such as zonula occludens-1 (ZO-1) and claudin-5, thereby preserving endothelial barrier structure[126]. They also improve endothelial cell survival under hypoxic conditions through activation of PI3K/AKT and MAPK signaling pathways[122].
AD: BBB repair and therapeutic delivery
In AD, BBB disruption contributes to disease progression by allowing peripheral inflammatory factors to enter the CNS. SC-EVs can mitigate this process. NSC-EVs have been reported to reduce BBB permeability and partially restore barrier function[127]. UCMSC-EVs suppress the TLR4/NF-κB signaling pathway via miR-125b-5p and provide vascular protection[128].
Critical appraisal and translational gaps
Although SC-EVs show vascular protective effects in preclinical studies, current models often rely on acute injury paradigms and simplified leakage assays. These approaches do not fully capture the complexity of chronic human vascular dysfunction. In particular, it remains uncertain whether SC-EVs can restore long-term neurovascular coupling and physiological transport across the BBB.
Regulation of pathological protein aggregation
The accumulation and propagation of misfolded proteins, including Aβ in AD and α-synuclein (α-Syn) in PD, are central to neurodegeneration. SC-EVs influence protein homeostasis through multiple mechanisms, including reducing protein production, enhancing degradation, and promoting clearance.
AD: clearance of Aβ via enzymatic degradation and phagocytosis
In AD, SC-EVs modulate several steps of the amyloid cascade. BMSC-EVs deliver miR-29c-3p, which suppresses β-secretase 1 (BACE1) and reduces the amyloidogenic APP processing[129]. They also promote Aβ clearance by activating the sphingosine kinase (SphK)/sphingosine-1-phosphate (S1P) pathway[114] and increasing the expression of amyloid-degrading enzymes such as neprilysin (NEP) and insulin-degrading enzyme (IDE)[113,130]. In addition, UCMSC-EVs enhance microglial phagocytosis of Aβ deposits, contributing to reduced plaque burden[92,115,131].
PD: attenuation of α-Syn aggregation and toxicity
In PD, SC-EVs reduce α-Syn accumulation and associated toxicity. Neural-differentiated ADMSC-EVs decrease intracellular α-Syn levels and may limit downstream neuroinflammatory responses[132]. NSC-EVs similarly reduce α-Syn burden in PD models[133]. These effects may alleviate lysosomal stress and help preserve dopaminergic neuron function.
Critical appraisal and translational gaps
Despite promising findings, current models often rely on artificial overexpression of mutant proteins, which does not fully reflect human disease progression. In addition, many studies focus on end-stage plaque measurements rather than dynamic processes such as oligomer formation and propagation. Therefore, it remains unclear whether SC-EVs can effectively halt long-term protein aggregation and disease progression in vivo.
Summary of therapeutic mechanisms: the pleiotropic synergy of SC-EVs
SC-EVs exert therapeutic effects through multiple coordinated mechanisms rather than a single target pathway. They deliver a combination of regulatory RNAs and proteins that influence inflammation, apoptosis, oxidative stress, synaptic function, and vascular integrity [Figure 3].
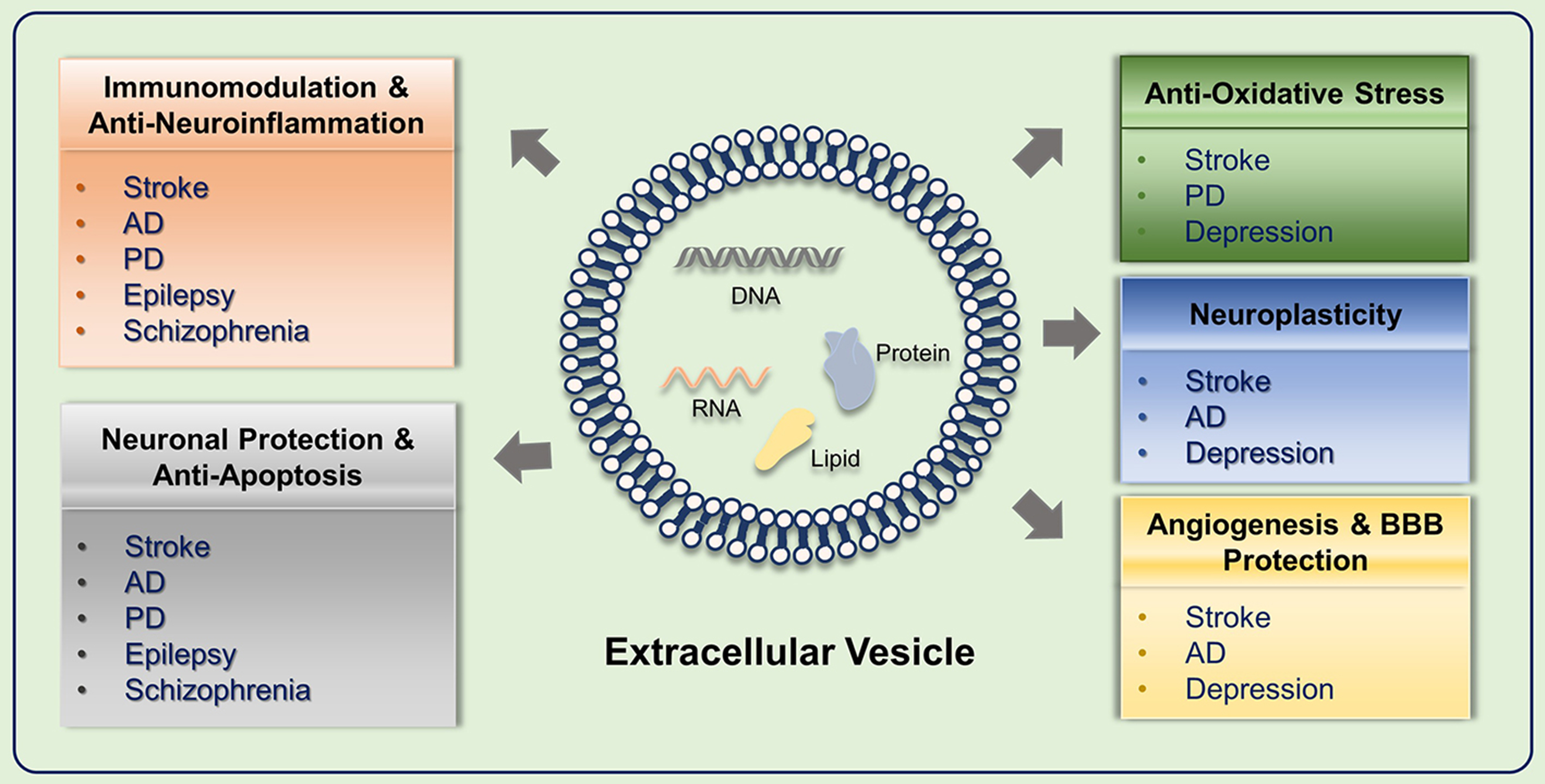
Figure 3. Integrated therapeutic mechanisms of SC-EVs in neurological disorders. This schematic illustrates six major functional pathways: (1) Regulation of Pathological Protein Aggregation to restore proteostasis in AD and PD; (2) Immunomodulation and Anti-Neuroinflammation alongside (3) Neuronal Protection and Anti-Apoptosis, which collectively provide broad cellular defense across acute injuries (stroke), chronic neurodegeneration (AD, PD), and psychiatric/seizure disorders (epilepsy, schizophrenia); (4) Anti-Oxidative Stress responses to restore intracellular redox balance in stroke, PD, and depression; (5) Neuroplasticity enhancement to rebuild synaptic circuitry; and (6) Angiogenesis and BBB Protection to repair the neurovascular unit in stroke and AD. SC-EVs: Stem cell-derived extracellular vesicles; AD: Alzheimer’s disease; PD: Parkinson’s disease; BBB: blood-brain barrier.
Unlike conventional pharmacological agents, EVs represent a cell-free system that retains key functional properties of stem cells while reducing safety concerns associated with cell transplantation. The molecular mediators underlying these effects are summarized in Table 2.
Summary of SC-EV therapeutic mechanisms and targets in neurological disorders
| Therapeutic mechanism | Cellular origin of SC-EVs | Key cargo | Target/Pathway | Disease type | Ref. |
| Immunomodulation and anti-neuroinflammation | BMSC | lncRNA ZFAS1 | miR-15a-5p | Stroke | [83] |
| BMSC | Data unavailable | NLRP3 | Stroke | [84] | |
| UCMSC | Data unavailable | MyD88/TRAF6/NF-κB | Depression | [105] | |
| ADMSC | Data unavailable | AMPK-mTOR | Depression | [106] | |
| UCMSC | Data unavailable | NRF2 | AD | [134] | |
| Oxidative stress regulation | NSC | Data unavailable | PINK1/Parkin | Stroke | [107] |
| ADMSC | miR-23b-3p | STAT1/GlyR1 | Epilepsy | [99] | |
| BMSC | miR-146a-5p | TRAF6 | Epilepsy | [135] | |
| UCMSC | NRF2, NLRP3 | NRF2/NF-κB/NLRP3 | Epilepsy | [101] | |
| Neuronal protection | BMSC | lncRNA KLF3-AS1 | miR-206/USP22 | Stroke | [85] |
| UCMSC | TSG-6 | STAT3-miR-7-NEDD4 | PD | [116] | |
| BMSC | Data unavailable | FTO siRNA; ATM mRNA | PD | [136] | |
| BMSC | Data unavailable | miR-26a | Depression | [108] | |
| BMSC | Data unavailable | BDNF | AD | [91] | |
| MSC | Data unavailable | BDNF | Schizophrenia | [104] | |
| Neurogenesis promotion | BMSC | Data unavailable | miR-21-5p | Stroke | [125] |
| UCMSC | Data unavailable | BDNF | Depression | [137] | |
| Synaptogenesis and neuroplasticity regulation | ADMSC | Data unavailable | Galectin-3 | Depression | [122] |
| BMSC | circRNA hsa_circ_0000288 | Caprin1 | Epilepsy | [117] | |
| Angiogenesis promotion | BMSC | Data unavailable | miR-126-3p; miR-140-5p; let-7c-5p | Stroke | [124] |
| Abnormal protein aggregation regulation | BMSC | Data unavailable | miR-29c-3p/BACE1; Wnt/β-catenin | AD | [129] |
| UCMSC | Data unavailable | NEP; miR-211-5p | AD | [131] | |
| BMSC | Data unavailable | IDE | AD | [113] | |
| BMSC | Data unavailable | SphK/S1P | AD | [114] |
Clinical translation of EV therapy in neurological disorders
SC-EVs have attracted increasing attention in clinical research. Although preclinical studies provide substantial mechanistic evidence, clinical translation remains at an early stage. Most trials are ongoing, and only a limited number have reported completed outcomes[138] [Table 3].
Overview of clinical trials involving SC-EVs for neurological disorders
| Disease/Condition | NCT Number | Sample size | Cellular origin of SC-EVs | Dosing regimen | Phase | Endpoints | Reported outcomes/Notes |
| Ischemic stroke | NCT06612710 | 69 | Human-induced neural stem cells | Patients received intravenous injection of EVs at a dosage of 4 × 109 particles/kg once daily for 7 days | Phase 1 | Assess the safety; MRI; Scale scores | Data unavailable |
| NCT07232563 | 9 | hUC-MSCs | Patients received hUC-MSC-sEV-001 via intranasal instillation at a dosage of 1 × 1011 particles per administration. The treatment was initiated on the day of enrollment and continued daily for 4 days | Phase 1 | Dose-Limiting Toxicity; Adverse events; Scale scores | Data unavailable | |
| Acute ischemic stroke | NCT06995625 | 18 | Wharton’s Jelly MSCs | EVs were administered once daily for 5 days. There were three cohorts, each with 3 to 6 participants, who received escalating doses: 4.8 × 1010, 9.6 × 1010, and 19.2 × 1010 particles, respectively | Phase 1 | Assess the safety; Maximum tolerated dose; Assess the efficacy | Evidence from non-human primate models underscored the potential for functional neurological restoration |
| NCT07145294 | 40 | iPSCs | EVs (2 mL, 2 × 1010 particles/mL) were administered intranasally twice weekly for 12 weeks, in combination with standard-of-care treatment for cerebrovascular disease | Phase 2 | Scale scores; Inflammatory indicators; Blood-brain barrier disruption indicators; Neuronal damage indicators | Data unavailable | |
| AD | NCT04388982 | 9 | Allogeneic adipose MSCs | Participants were assigned to three dosage groups: low-dose (5 μg), medium-dose (10 μg), and high-dose (20 μg). All groups received intranasal instillations (1 mL) twice weekly for 12 weeks | Phase 2 | Assess the safety; Cognitive function; Quality of life evaluation; MRI Neuroimaging; Changes of AD biomarkers | The treatment was found to be safe and well-tolerated. Notably, the medium-dose group showed potential cognitive improvements and reduced hippocampal volume loss |
| PD | NCT05152394 | 20 | Allogeneic adult umbilical cord MSCs | Participants received two doses of EVs via intranasal instillation on consecutive days. Each dose consisted of 4 mL of solution containing approximately 8 × 1011 exosomes | Phase 1 | Assess the safety | Data unavailable |
| Refractory focal epilepsy | NCT05886205 | 34 | iPSCs | Participants were assigned to four cohorts: a low-dose group (2 μg, n = 8), a mid-dose group (6 μg, n = 8), a high-dose group (18 μg, n = 8), and a dose-expansion group (n = 10). All participants received iPSC-Exos (200 μL) via nasal drip twice daily for 12 weeks | Early phase 1 | Assess the safety; Seizure frequency | Data unavailable |
| Neurodegenerative diseases | NCT06607900 | 120 | Human umbilical cord MSCs | Intranasal administration | Phase 1 | Scale scores | Data unavailable |
A completed clinical trial (NCT04388982) evaluated intranasal administration of allogeneic adipose-derived MSC-EVs (ahaMSC-EVs) in patients with mild-to-moderate AD. Participants received treatment twice weekly for 12 weeks across three dose groups. The intranasal route is of particular interest because it enables delivery to the CNS via olfactory and trigeminal pathways, thereby bypassing the BBB. The study demonstrated that repeated administration of ahaMSC-EVs was safe and well tolerated. The medium-dose group showed greater improvement in cognitive function and reduced hippocampal atrophy compared with the other groups[139]. These findings suggest that EV efficacy may not increase linearly with dose, and they highlight the need for optimized dosing strategies and pharmacokinetic characterization in future studies.
Challenges and bottlenecks in clinical translation
Despite the recognized advantages of SC-EVs, including low immunogenicity and the ability to cross the BBB, their clinical translation remains limited by several challenges. These include technical constraints in production, regulatory complexity, and economic considerations.
Production standardization and scalability
A major barrier to clinical application is the lack of standardized Good Manufacturing Practice (GMP)-compliant protocols for EV production and characterization. EVs are highly sensitive to the cellular microenvironment; therefore, variations in stem cell source, passage number, and culture conditions (e.g., bioreactor shear stress) can alter EV composition and subpopulation profiles. This variability contributes to inconsistencies in bioactive cargo, including miRNAs and proteins, and compromises reproducibility across batches. In addition, despite the development of MISEV guidelines, standardized and high-throughput methods for particle quantification and surface marker characterization (e.g., CD63, CD81) are still lacking, which further limits comparability between studies[17,140].
Dosage optimization and efficacy validation
Defining the clinically relevant dosing strategy remains a key pharmacokinetic challenge. Current preclinical studies use heterogeneous dosing regimens, and EVs do not follow conventional small-molecule pharmacokinetics. As a result, dose extrapolation from animal models to humans using standard allometric scaling is unreliable. Moreover, most clinical studies remain exploratory, with limited sample sizes, heterogeneous endpoints, and insufficient long-term follow-up, making it difficult to assess the sustained therapeutic effects[141,142].
Long-term safety and pharmacokinetics
Long-term biodistribution and safety data for SC-EVs remain limited. Although EVs are thought to preferentially accumulate at sites of injury or inflammation, systemically administered EVs are rapidly cleared by the mononuclear phagocyte system. A substantial proportion accumulates in off-target organs, particularly the liver and spleen. The long-term consequences of this peripheral accumulation, as well as the potential for immune responses following repeated administration, have not yet been fully characterized and require further investigation[143-145].
Regulatory and economic constraints
The regulatory framework for EV-based therapies is still evolving and varies across regions. Differences in classification criteria between agencies such as the US FDA and the European Medicines Agency (EMA) can lead to divergent approval pathways, particularly for engineered or modified EV products[146,147]. In addition, production costs remain high due to the need for controlled cell culture systems, advanced purification technologies, and comprehensive quality control. Current estimates suggest that EV-based therapies are substantially more expensive than conventional pharmaceuticals[148,149]. However, comprehensive health economic evaluations are still lacking.
These challenges are interrelated and will require coordinated efforts to address. Advances in scalable production technologies, improved standardization, and greater regulatory alignment are needed. Ultimately, well-designed, large-scale clinical trials will be necessary to establish the safety and efficacy of SC-EV-based therapies.
CONCLUSION
SC-EVs represent a promising approach for the treatment of neurological disorders, owing to their ability to cross the BBB, deliver bioactive cargo, and modulate multiple pathological pathways. Future research should prioritize improving targeting efficiency, optimizing cargo loading, and evaluating combination approaches with existing therapies. EV-associated molecules may also serve as valuable biomarkers for disease monitoring and treatment response[150]. To support clinical translation, standard manufacturing protocols, well-defined dosing strategies, and reliable delivery systems must be established. Addressing these issues will be essential for moving SC-EVs-based therapies from experimental models toward clinical application.
DECLARATIONS
Authors’ contributions
Made substantial contributions to conception and review & editing and design of the study: Ma C, Sun C
Performed original draft, investigation, software and data curation: Fan H, Wang S
Performed original draft and data curation: Li Z, Yu T
Availability of data and materials
Not applicable.
AI and AI-assisted tools statement
Not applicable.
Financial support and sponsorship
This work was supported partly by grants from the National Natural Science Foundation of China (32260194 and 32560199), the Natural Science Foundation of Jiangxi Province (20224BAB206042, 20224BAB206038 and 20252BAC250141).
Conflicts of interest
All authors declared that there are no conflicts of interest.
Ethical approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Copyright
© The Author(s) 2026.
REFERENCES
1. World Health Organization. Global status report on neurology. Available from: https://www.who.int/publications/i/item/9789240116139. [Last accessed on 22 May 2026].
2. Feigin VL, Vos T, Nichols E, et al. The global burden of neurological disorders: translating evidence into policy. Lancet Neurol. 2020;19:255-65.
3. Furtado D, Björnmalm M, Ayton S, Bush AI, Kempe K, Caruso F. Overcoming the blood-brain barrier: the role of nanomaterials in treating neurological diseases. Adv Mater. 2018;30:e1801362.
4. Ying C, Zhang J, Zhang H, et al. Stem cells in central nervous system diseases: promising therapeutic strategies. Exp Neurol. 2023;369:114543.
5. Arbade G, Jose JV, Gulbake A, Kadam S, Kashte SB. From stem cells to extracellular vesicles: a new horizon in tissue engineering and regenerative medicine. Cytotechnology. 2024;76:363-401.
6. Zhang X, Che X, Zhang S, et al. Mesenchymal stem cell-derived extracellular vesicles for human diseases. Extracell Vesicles Circ Nucl Acids. 2024;5:64-82.
7. Yin T, Liu Y, He B, et al. Cell primitive-based biomimetic nanomaterials for Alzheimer’s disease targeting and therapy. Mater Today Bio. 2023;22:100789.
9. Choudhery MS, Arif T, Mahmood R, Harris DT. Stem cell-based acellular therapy: insight into biogenesis, bioengineering and therapeutic applications of exosomes. Biomolecules. 2024;14:792.
10. Shi M, Chu F, Zhu J. Stem cells therapy in neurodegenerative and neuroimmune diseases: current status of treatments and future prospects. Pharmacol Res. 2025;221:107960.
11. Yu S, Yu S, Liu H, Liao N, Liu X. Enhancing mesenchymal stem cell survival and homing capability to improve cell engraftment efficacy for liver diseases. Stem Cell Res Ther. 2023;14:235.
12. Su Z, Dong H, Fang X, Zhang W, Duan H. Frontier progress and translational challenges of pluripotent differentiation of stem cells. Front Genet. 2025;16:1583391.
13. Stahl PD, Raposo G. Extracellular vesicles: exosomes and microvesicles, integrators of homeostasis. Physiology. 2019;34:169-77.
15. Wang Y, Wang H, Tan J, et al. Therapeutic effect of mesenchymal stem cells and their derived exosomes in diseases. Mol Biomed. 2025;6:34.
16. Théry C, Witwer KW, Aikawa E, et al. Minimal information for studies of extracellular vesicles 2018 (MISEV2018): a position statement of the International Society for Extracellular Vesicles and update of the MISEV2014 guidelines. J Extracell Vesicles. 2018;7:1535750.
17. Welsh JA, Goberdhan DCI, O’Driscoll L, et al. ; MISEV Consortium. Minimal information for studies of extracellular vesicles (MISEV2023): from basic to advanced approaches. J Extracell Vesicles. 2024;13:e12404.
18. van Griensven M, Balmayor ER. Extracellular vesicles are key players in mesenchymal stem cells’ dual potential to regenerate and modulate the immune system. Adv Drug Deliv Rev. 2024;207:115203.
19. Pfrieger FW, Vitale N. Cholesterol and the journey of extracellular vesicles. J Lipid Res. 2018;59:2255-61.
20. Guevara-Olaya L, Chimal-Vega B, Castañeda-Sánchez CY, et al. LDL promotes disorders in β-cell cholesterol metabolism, implications on insulin cellular communication mediated by EVs. Metabolites. 2022;12:754.
21. Longo A, Manganelli V, Misasi R, et al. Extracellular vesicles in the crosstalk of autophagy and apoptosis: a role for lipid rafts. Cells. 2025;14:749.
22. Kalluri R, LeBleu VS. The biology, function, and biomedical applications of exosomes. Science. 2020;367:eaau6977.
23. Rädler J, Gupta D, Zickler A, Andaloussi SE. Exploiting the biogenesis of extracellular vesicles for bioengineering and therapeutic cargo loading. Mol Ther. 2023;31:1231-50.
24. Gurung S, Perocheau D, Touramanidou L, Baruteau J. The exosome journey: from biogenesis to uptake and intracellular signalling. Cell Commun Signal. 2021;19:47.
25. Padinharayil H, Varghese J, Wilson C, George A. Mesenchymal stem cell-derived exosomes: characteristics and applications in disease pathology and management. Life Sci. 2024;342:122542.
26. Rather HA, Almousa S, Craft S, Deep G. Therapeutic efficacy and promise of stem cell-derived extracellular vesicles in Alzheimer’s disease and other aging-related disorders. Ageing Res Rev. 2023;92:102088.
27. Sawicki S, Bugno-Poniewierska M, Żurowski J, et al. Comparative transcriptome and microRNA profiles of equine mesenchymal stem cells, fibroblasts, and their extracellular vesicles. Genes. 2025;16:936.
28. Rezaie J, Nejati V, Mahmoodi M, Ahmadi M. Mesenchymal stem cells derived extracellular vesicles: a promising nanomedicine for drug delivery system. Biochem Pharmacol. 2022;203:115167.
29. Yuan X, Sun L, Jeske R, et al. Engineering extracellular vesicles by three-dimensional dynamic culture of human mesenchymal stem cells. J Extracell Vesicles. 2022;11:e12235.
30. Bertolino GM, Maumus M, Jorgensen C, Noël D. Therapeutic potential in rheumatic diseases of extracellular vesicles derived from mesenchymal stromal cells. Nat Rev Rheumatol. 2023;19:682-94.
31. Xiao Y, Hoorain I, Zhang L, et al. Unpacking the anti-fibrotic arsenal: molecular mechanisms and therapeutic translation of MSC-derived exosomes in pulmonary fibrosis. Front Immunol. 2025;16:1725041.
32. Yoo D, Jung SY, Go D, et al. Functionalized extracellular vesicles of mesenchymal stem cells for regenerative medicine. J Nanobiotechnology. 2025;23:219.
33. Li F, Zhang J, Chen A, et al. Combined transplantation of neural stem cells and bone marrow mesenchymal stem cells promotes neuronal cell survival to alleviate brain damage after cardiac arrest via microRNA-133b incorporated in extracellular vesicles. Aging. 2021;13:262-78.
34. Tan Q, Xia D, Ying X. miR-29a in exosomes from bone marrow mesenchymal stem cells inhibit fibrosis during endometrial repair of intrauterine adhesion. Int J Stem Cells. 2020;13:414-23.
35. Wei R, Zhang L, Hu W, Shang X, He Y, Zhang W. Zeb2/axin2-enriched BMSC-derived exosomes promote post-stroke functional recovery by enhancing neurogenesis and neural plasticity. J Mol Neurosci. 2022;72:69-81.
36. Ding Z, Greenberg ZF, Serafim MF, et al. Understanding molecular characteristics of extracellular vesicles derived from different types of mesenchymal stem cells for therapeutic translation. Extracell Vesicle. 2024;3:100034.
37. Abdal Dayem A, Lee SB, Kim K, et al. Production of mesenchymal stem cells through stem cell reprogramming. Int J Mol Sci. 2019;20:1922.
38. Yamanaka S. Pluripotent stem cell-based cell therapy-promise and challenges. Cell Stem Cell. 2020;27:523-31.
39. Shi Y, Yang X, Wang S, et al. Human umbilical cord mesenchymal stromal cell-derived exosomes protect against MCD-induced NASH in a mouse model. Stem Cell Res Ther. 2022;13:517.
40. Zhang Z, Mi T, Jin L, et al. Comprehensive proteomic analysis of exosome mimetic vesicles and exosomes derived from human umbilical cord mesenchymal stem cells. Stem Cell Res Ther. 2022;13:312.
41. Yaghoubi Y, Movassaghpour A, Zamani M, Talebi M, Mehdizadeh A, Yousefi M. Human umbilical cord mesenchymal stem cells derived-exosomes in diseases treatment. Life Sci. 2019;233:116733.
42. Mazini L, Rochette L, Amine M, Malka G. Regenerative capacity of adipose derived stem cells (ADSCs), comparison with mesenchymal stem cells (MSCs). Int J Mol Sci. 2019;20:2523.
43. Nashwan S, Ismail MA, Saleh T, et al. Comparative analysis of extracellular vesicles from induced and adipose-derived mesenchymal stem cells: implications for regenerative medicine. PLoS One. 2025;20:e0325065.
44. Heo JS, Choi Y, Kim HO. Adipose-derived mesenchymal stem cells promote M2 macrophage phenotype through exosomes. Stem Cells Int. 2019;2019:7921760.
45. Xing X, Han S, Cheng G, Ni Y, Li Z, Li Z. Proteomic analysis of exosomes from adipose-derived mesenchymal stem cells: a novel therapeutic strategy for tissue injury. Biomed Res Int. 2020;2020:6094562.
46. Hong P, Yang H, Wu Y, Li K, Tang Z. The functions and clinical application potential of exosomes derived from adipose mesenchymal stem cells: a comprehensive review. Stem Cell Res Ther. 2019;10:242.
47. Tian L, Zhang D, Wang Z, et al. MiR-877, an exosomal miRNA from mechanical stretch induced adipose derived stromal cells, enhances fracture healing in nonunion rats with type 2 diabetes mellitus. Stem Cell Res Ther. 2025;16:575.
48. Chang LH, Wu SC, Chen CH, et al. Exosomes derived from hypoxia-cultured human adipose stem cells alleviate articular chondrocyte inflammaging and post-traumatic osteoarthritis progression. Int J Mol Sci. 2023;24:13414.
49. Belair C, Sim S, Kim KY, Tanaka Y, Park IH, Wolin SL. The RNA exosome nuclease complex regulates human embryonic stem cell differentiation. J Cell Biol. 2019;218:2564-82.
50. Kwon MH, Song KM, Limanjaya A, et al. Embryonic stem cell-derived extracellular vesicle-mimetic nanovesicles rescue erectile function by enhancing penile neurovascular regeneration in the streptozotocin-induced diabetic mouse. Sci Rep. 2019;9:20072.
51. Cruz L, Arevalo Romero JA, Brandão Prado M, Santos TG, Hohmuth Lopes M. Evidence of extracellular vesicles biogenesis and release in mouse embryonic stem cells. Stem Cell Rev Rep. 2018;14:262-76.
52. Jeske R, Bejoy J, Marzano M, Li Y. Human pluripotent stem cell-derived extracellular vesicles: characteristics and applications. Tissue Eng Part B Rev. 2020;26:129-44.
53. Liu M, Qiu Y, Xue Z, et al. Small extracellular vesicles derived from embryonic stem cells restore ovarian function of premature ovarian failure through PI3K/AKT signaling pathway. Stem Cell Res Ther. 2020;11:3.
54. Wang AYL, Kao HK, Liu YY, Loh CYY. Engineered extracellular vesicles derived from pluripotent stem cells: a cell-free approach to regenerative medicine. Burns Trauma. 2025;13:tkaf013.
55. Fan C, Zhang E, Joshi J, Yang J, Zhang J, Zhu W. Utilization of human induced pluripotent stem cells for cardiac repair. Front Cell Dev Biol. 2020;8:36.
56. Barreca MM, Cancemi P, Geraci F. Mesenchymal and induced pluripotent stem cells-derived extracellular vesicles: the new frontier for regenerative medicine? Cells. 2020;9:1163.
57. Xu M, Chen G, Dong Y, et al. Stable expression of a truncated TLX variant drives differentiation of induced pluripotent stem cells into self-renewing neural stem cells for production of extracellular vesicles. Stem Cell Res Ther. 2022;13:436.
58. Wang Y, Zhao J, Guo Q, et al. Neural stem cell-derived exosomes improve neurite outgrowth and cognitive function through transferring miR-132-3p. Exp Neurol. 2025;388:115224.
59. Yuan P, Ding L, Chen H, et al. Neural stem cell-derived exosomes regulate neural stem cell differentiation through miR-9-Hes1 axis. Front Cell Dev Biol. 2021;9:601600.
60. Derkus B, Isik M, Eylem CC, et al. Xenogenic neural stem cell-derived extracellular nanovesicles modulate human mesenchymal stem cell fate and reconstruct metabolomic structure. Adv Biol. 2022;6:e2101317.
61. Luo H, Ye G, Liu Y, et al. miR-150-3p enhances neuroprotective effects of neural stem cell exosomes after hypoxic-ischemic brain injury by targeting CASP2. Neurosci Lett. 2022;779:136635.
62. Ma K, Xu H, Zhang J, et al. Insulin-like growth factor-1 enhances neuroprotective effects of neural stem cell exosomes after spinal cord injury via an miR-219a-2-3p/YY1 mechanism. Aging. 2019;11:12278-94.
63. Doktor F, Figueira RL, Fortuna V, et al. Amniotic fluid stem cell extracellular vesicles promote lung development via TGF-beta modulation in a fetal rat model of oligohydramnios. J Control Release. 2025;377:427-41.
64. Kim T, Hong JW, Lee LP. Efficient methods of isolation and purification of extracellular vesicles. Nano Converg. 2025;12:45.
65. Li K, Guo B, Gu J, et al. Emerging advances in drug delivery systems (DDSs) for optimizing cancer complications. Mater Today Bio. 2025;30:101375.
66. Ratushnyak MG, Shaposhnikova DA, Gubani DF, Kondratyev KV. Characterization of stem cell exosomes isolated by ultracentrifugation, depth filtration, and precipitation with polyethylene glycol. Nanotechnologies Russ. 2025;20:78-87.
67. Khan MI, Chen K, Zhang L, et al. Efficient purification of exosomes and subpopulations in fresh bovine milk via fine isolation of free-flow isoelectric focusing in large scale. CEJ. 2025;508:160850.
68. Chen S, Shiesh SC, Lee GB, Chen C. Two-step magnetic bead-based (2MBB) techniques for immunocapture of extracellular vesicles and quantification of microRNAs for cardiovascular diseases: a pilot study. PLoS One. 2020;15:e0229610.
69. Kverneland AH, Østergaard O, Emdal KB, Svane IM, Olsen JV. Differential ultracentrifugation enables deep plasma proteomics through enrichment of extracellular vesicles. Proteomics. 2023;23:e2200039.
70. Benmoussa A, Michel S, Gilbert C, Provost P. Isolating multiple extracellular vesicles subsets, including exosomes and membrane vesicles, from bovine milk using sodium citrate and differential ultracentrifugation. Bio Protoc. 2020;10:e3636.
71. Nielsen T, Kristensen AF, Pedersen S, Christiansen G, Kristensen SR. Investigation of procoagulant activity in extracellular vesicles isolated by differential ultracentrifugation. J Extracell Vesicles. 2018;7:1454777.
72. Langevin SM, Kuhnell D, Orr-Asman MA, et al. Balancing yield, purity and practicality: a modified differential ultracentrifugation protocol for efficient isolation of small extracellular vesicles from human serum. RNA Biol. 2019;16:5-12.
73. Musante L, Tataruch-Weinert D, Kerjaschki D, Henry M, Meleady P, Holthofer H. Residual urinary extracellular vesicles in ultracentrifugation supernatants after hydrostatic filtration dialysis enrichment. J Extracell Vesicles. 2017;6:1267896.
74. Soares M, Pinto MM, Nobre RJ, et al. Isolation of extracellular vesicles from human follicular fluid: size-exclusion chromatography versus ultracentrifugation. Biomolecules. 2023;13:278.
75. Foers AD, Chatfield S, Dagley LF, et al. Enrichment of extracellular vesicles from human synovial fluid using size exclusion chromatography. J Extracell Vesicles. 2018;7:1490145.
76. Böing AN, van der Pol E, Grootemaat AE, Coumans FA, Sturk A, Nieuwland R. Single-step isolation of extracellular vesicles by size-exclusion chromatography. J Extracell Vesicles. 2014;3:23430.
77. Jung JH, Back W, Yoon J, et al. Dual size-exclusion chromatography for efficient isolation of extracellular vesicles from bone marrow derived human plasma. Sci Rep. 2021;11:217.
78. Singh M, Tiwari PK, Kashyap V, Kumar S. Proteomics of extracellular vesicles: recent updates, challenges and limitations. Proteomes. 2025;13:12.
79. Yang D, Zhang W, Zhang H, et al. Progress, opportunity, and perspective on exosome isolation - efforts for efficient exosome-based theranostics. Theranostics. 2020;10:3684-707.
80. Duong P, Chung A, Bouchareychas L, Raffai RL. Cushioned-density gradient ultracentrifugation (C-DGUC) improves the isolation efficiency of extracellular vesicles. PLoS One. 2019;14:e0215324.
81. Parimon T, Garrett NE 3rd, Chen P, Antes TJ. Isolation of extracellular vesicles from murine bronchoalveolar lavage fluid using an ultrafiltration centrifugation technique. J Vis Exp. 2018;141:e58310.
82. Fortunato D, Giannoukakos S, Giménez-Capitán A, Hackenberg M, Molina-Vila MA, Zarovni N. Selective isolation of extracellular vesicles from minimally processed human plasma as a translational strategy for liquid biopsies. Biomark Res. 2022;10:57.
83. Yang H, Chen J. Bone marrow mesenchymal stem cell-derived exosomes carrying long noncoding RNA ZFAS1 alleviate oxidative stress and inflammation in ischemic stroke by inhibiting microRNA-15a-5p. Metab Brain Dis. 2022;37:2545-57.
84. Liu X, Zhang M, Liu H, et al. Bone marrow mesenchymal stem cell-derived exosomes attenuate cerebral ischemia-reperfusion injury-induced neuroinflammation and pyroptosis by modulating microglia M1/M2 phenotypes. Exp Neurol. 2021;341:113700.
85. Xie X, Cao Y, Dai L, Zhou D. Bone marrow mesenchymal stem cell-derived exosomal lncRNA KLF3-AS1 stabilizes Sirt1 protein to improve cerebral ischemia/reperfusion injury via miR-206/USP22 axis. Mol Med. 2023;29:3.
86. Zhang Z, Ji R, Liu Z, et al. hUMSC-Exosomes suppress TREM1-p38 MAPK signaling via HMGB1-dependent mechanisms to reprogram microglial function and promote neuroprotection in ischemic stroke. J Nanobiotechnology. 2025;23:572.
87. Liu Q, Ma H, Liao J, et al. MiR-125a-5p in extracellular vesicles of neural stem cells acts as a crosstalk signal modulating neuroinflammatory microenvironment to alleviate cerebral ischemia-reperfusion injury. Theranostics. 2025;15:7064-89.
88. Wang J, Zhao M, Fu D, et al. Human neural stem cell-derived extracellular vesicles protect against ischemic stroke by activating the PI3K/AKT/mTOR pathway. Neural Regen Res. 2025;20:3245-58.
89. Wang QS, Xiao RJ, Peng J, Yu ZT, Fu JQ, Xia Y. Bone marrow mesenchymal stem cell-derived exosomal KLF4 alleviated ischemic stroke through inhibiting N6-methyladenosine modification level of Drp1 by targeting lncRNA-ZFAS1. Mol Neurobiol. 2023;60:3945-62.
90. Cui GH, Guo HD, Li H, et al. RVG-modified exosomes derived from mesenchymal stem cells rescue memory deficits by regulating inflammatory responses in a mouse model of Alzheimer’s disease. Immun Ageing. 2019;16:10.
91. Liu S, Fan M, Xu JX, et al. Exosomes derived from bone-marrow mesenchymal stem cells alleviate cognitive decline in AD-like mice by improving BDNF-related neuropathology. J Neuroinflammation. 2022;19:35.
92. Ding M, Shen Y, Wang P, et al. Exosomes isolated from human umbilical cord mesenchymal stem cells alleviate neuroinflammation and reduce amyloid-beta deposition by modulating microglial activation in Alzheimer’s disease. Neurochem Res. 2018;43:2165-77.
93. Losurdo M, Pedrazzoli M, D’Agostino C, et al. Intranasal delivery of mesenchymal stem cell-derived extracellular vesicles exerts immunomodulatory and neuroprotective effects in a 3xTg model of Alzheimer’s disease. Stem Cells Transl Med. 2020;9:1068-84.
94. Li B, Chen Y, Zhou Y, et al. Neural stem cell-derived exosomes promote mitochondrial biogenesis and restore abnormal protein distribution in a mouse model of Alzheimer’s disease. Neural Regen Res. 2024;19:1593-601.
95. Zhang L, Zhan M, Sun H, et al. Mesenchymal stem-cell-derived exosomes loaded with phosphorus dendrimers and quercetin treat Parkinson’s disease by modulating inflammatory immune microenvironment. ACS Appl Mater Interfaces. 2025;17:32013-27.
96. Cai Y, Zhang MM, Wang M, Jiang ZH, Tan ZG. Bone marrow-derived mesenchymal stem cell-derived exosomes containing gli1 alleviate microglial activation and neuronal apoptosis in vitro and in a mouse Parkinson disease model by direct inhibition of Sp1 signaling. J Neuropathol Exp Neurol. 2022;81:522-34.
97. Zhang ZX, Zhou YJ, Gu P, et al. Exosomes derived from human umbilical cord mesenchymal stem cells alleviate Parkinson’s disease and neuronal damage through inhibition of microglia. Neural Regen Res. 2023;18:2291-300.
98. Lee EJ, Choi Y, Lee HJ, Hwang DW, Lee DS. Human neural stem cell-derived extracellular vesicles protect against Parkinson’s disease pathologies. J Nanobiotechnology. 2022;20:198.
99. Yang X, Yang X, Sun A, Chen S, Wang X, Zhao X. The miR-23b-3p from adipose-derived stem cell exosomes alleviate inflammation in mice experiencing kainic acid-induced epileptic seizures. Neuroreport. 2024;35:612-20.
100. Derisfard F, Jafarinezhad Z, Azarpira N, Namavar MR, Aligholi H. Exosomes obtained from human adipose-derived stem cells alleviate epileptogenesis in the pentylenetetrazol model of epilepsy. Neuroreport. 2025;36:161-8.
101. Che J, Wang H, Dong J, et al. Human umbilical cord mesenchymal stem cell-derived exosomes attenuate neuroinflammation and oxidative stress through the NRF2/NF-κB/NLRP3 pathway. CNS Neurosci Ther. 2024;30:e14454.
102. Niu Y, Wang X, Li M, Niu B. Exosomes from human umbilical cord Mesenchymal stem cells attenuates stress-induced hippocampal dysfunctions. Metab Brain Dis. 2020;35:1329-40.
103. Ma L, Xiao C, Zhang Z, Zhan YA. Exosomes secreted from induced pluripotent stem cell ameliorate the lipopolysaccharide induced neuroinflammatory response via lncRNA-0949. Immun Inflamm Dis. 2024;12:e1155.
104. Zhong XL, Huang Y, Du Y, et al. Unlocking the therapeutic potential of exosomes derived from nasal olfactory mucosal mesenchymal stem cells: restoring synaptic plasticity, neurogenesis, and neuroinflammation in schizophrenia. Schizophr Bull. 2024;50:600-14.
105. Li P, Zhang F, Huang C, et al. Exosomes derived from DPA-treated UCMSCs attenuated depression-like behaviors and neuroinflammation in a model of depression induced by chronic stress. J Neuroimmune Pharmacol. 2024;19:55.
106. Sun S, Rong J, Wang C, et al. Intranasal administration of exosomes derived from adipose mesenchymal stem cells ameliorates depressive-like behaviors and inhibits inflammation via AMPK/mTOR-mediated autophagy. J Affect Disord. 2025;382:227-47.
107. Zhao M, Wang J, Zhu S, et al. Human neural stem cell-derived exosomes activate PINK1/Parkin pathway to protect against oxidative stress-induced neuronal injury in ischemic stroke. J Transl Med. 2025;23:402.
108. Guo H, Huang B, Wang Y, Zhang Y, Ma Q, Ren Y. Bone marrow mesenchymal stem cells-derived exosomes improve injury of hippocampal neurons in rats with depression by upregulating microRNA-26a expression. Int Immunopharmacol. 2020;82:106285.
109. Abdel-Gawad DRI, Khalil F, Shehata O, et al. Role of bone marrow mesenchymal stem cell-derived exosomes in reducing neurotoxicity and depression-like behaviors induced by doxorubicin in rats. Toxicol Res. 2024;13:tfae159.
110. Kuang Y, Zheng X, Zhang L, et al. Adipose-derived mesenchymal stem cells reduce autophagy in stroke mice by extracellular vesicle transfer of miR-25. J Extracell Vesicles. 2020;10:e12024.
111. Wang Y, Niu H, Li L, et al. Anti-CHAC1 exosomes for nose-to-brain delivery of miR-760-3p in cerebral ischemia/reperfusion injury mice inhibiting neuron ferroptosis. J Nanobiotechnology. 2023;21:109.
112. Ye YC, Chang ZH, Wang P, et al. Infarct-preconditioning exosomes of umbilical cord mesenchymal stem cells promoted vascular remodeling and neurological recovery after stroke in rats. Stem Cell Res Ther. 2022;13:378.
113. Ebrahim N, Al Saihati HA, Alali Z, et al. Exploring the molecular mechanisms of MSC-derived exosomes in Alzheimer’s disease: autophagy, insulin and the PI3K/Akt/mTOR signaling pathway. Biomed Pharmacother. 2024;176:116836.
114. Wang X, Yang G. Bone marrow mesenchymal stem cells-derived exosomes reduce Aβ deposition and improve cognitive function recovery in mice with Alzheimer’s disease by activating sphingosine kinase/sphingosine-1-phosphate signaling pathway. Cell Biol Int. 2021;45:775-84.
115. Chen YA, Lu CH, Ke CC, et al. Mesenchymal stem cell-derived exosomes ameliorate Alzheimer's disease pathology and improve cognitive deficits. Biomedicines. 2021;9:594.
116. Huang D, Zhang M, Tan Z. Bone marrow stem cell-exo-derived TSG-6 attenuates 1-methyl-4-phenylpyridinium+-induced neurotoxicity via the STAT3/miR-7/NEDD4/LRRK2 axis. J Neuropathol Exp Neurol. 2022;81:621-34.
117. Guo L, Lv N, Ji JL, et al. Circular RNA hsa_circ_0000288 protects against epilepsy in mice by binding to and stabilizing caprin1 protein. Acta Pharmacol Sin. 2025;46:1592-609.
118. Nazari S, Pourmand SM, Motevaseli E, Hassanzadeh G. Mesenchymal stem cells (MSCs) and MSC-derived exosomes in animal models of central nervous system diseases: Targeting the NLRP3 inflammasome. IUBMB Life. 2023;75:794-810.
119. Tsivion-Visbord H, Perets N, Sofer T, et al. Mesenchymal stem cells derived extracellular vesicles improve behavioral and biochemical deficits in a phencyclidine model of schizophrenia. Transl Psychiatry. 2020;10:305.
120. Liu H, Yan XJ, Hu JL, Pan H, Mao X, Cheng Y. Exosome derived from bone marrow derived mesenchymal stem cells prevents LPS-induced depressive like behaviors. Brain Res Bull. 2025;231:111527.
121. Upadhya R, Madhu LN, Attaluri S, et al. Extracellular vesicles from human iPSC-derived neural stem cells: miRNA and protein signatures, and anti-inflammatory and neurogenic properties. J Extracell Vesicles. 2020;9:1809064.
122. Fu W, Guo Y, Wu P, et al. hADMSC-Evs attenuates depressive and anxiety - like behaviors in chronic liver disease via suppressing Liver-Brain Galectin3 signaling. Brain Behav Immun. 2025;130:106082.
123. Lin SL, Chang YW, Lee W, et al. Role of STAT3-FOXO3 signaling in the modulation of neuroplasticity by PD-L1-HGF-decorated mesenchymal stem cell-derived exosomes in a murine stroke model. Adv Sci. 2024;11:e2404882.
124. Gregorius J, Wang C, Stambouli O, et al. Small extracellular vesicles obtained from hypoxic mesenchymal stromal cells have unique characteristics that promote cerebral angiogenesis, brain remodeling and neurological recovery after focal cerebral ischemia in mice. Basic Res Cardiol. 2021;116:40.
125. Hu H, Hu X, Li L, et al. Exosomes derived from bone marrow mesenchymal stem cells promote angiogenesis in ischemic stroke mice via upregulation of miR-21-5p. Biomolecules. 2022;12:883.
126. Li Y, Liu B, Zhao T, et al. Comparative study of extracellular vesicles derived from mesenchymal stem cells and brain endothelial cells attenuating blood-brain barrier permeability via regulating Caveolin-1-dependent ZO-1 and Claudin-5 endocytosis in acute ischemic stroke. J Nanobiotechnology. 2023;21:70.
127. Liu Y, Huber CC, Wang H. Disrupted blood-brain barrier in 5×FAD mouse model of Alzheimer’s disease can be mimicked and repaired in vitro with neural stem cell-derived exosomes. Biochem Biophys Res Commun. 2020;525:192-6.
128. Qiu L, Cai Y, Geng Y, et al. Mesenchymal stem cell-derived extracellular vesicles attenuate tPA-induced blood-brain barrier disruption in murine ischemic stroke models. Acta Biomater. 2022;154:424-42.
129. Sha S, Shen X, Cao Y, Qu L. Mesenchymal stem cells-derived extracellular vesicles ameliorate Alzheimer's disease in rat models via the microRNA-29c-3p/BACE1 axis and the Wnt/β-catenin pathway. Aging. 2021;13:15285-306.
130. Elia CA, Tamborini M, Rasile M, et al. Intracerebral injection of extracellular vesicles from mesenchymal stem cells exerts reduced Aβ plaque burden in early stages of a preclinical model of Alzheimer’s disease. Cells. 2019;8:1059.
131. Chen H, Huang Z, Lei A, Yu X, Shen M, Wu D. miRNA-211-5p inhibition enhances the protective effect of hucMSC-derived exosome in Aβ1-40 -induced SH-SY5Y cells by increasing NEP expression. J Biochem Mol Toxicol. 2024;38:e23624.
132. Ramalingam M, Jang S, Hwang J, Cho HH, Kim BC, Jeong HS. Neural-induced human adipose tissue-derived stem cell secretome exerts neuroprotection against rotenone-induced Parkinson's disease in rats. Stem Cell Res Ther. 2025;16:193.
133. Guo X, Xing J, Shi X, Cao Y, An L. Effects of neural stem cell-derived exosomes on MPTP-induced Parkinson’s disease in mice. Naturwissenschaften. 2025;112:53.
134. Wang H, Liu Y, Li J, et al. Tail-vein injection of MSC-derived small extracellular vesicles facilitates the restoration of hippocampal neuronal morphology and function in APP / PS1 mice. Cell Death Discov. 2021;7:230.
135. Huang X, Liu X, Zeng J, Du P, Huang X, Lin J. Bone marrow mesenchymal stem cell-derived exosomal microRNA regulates microglial polarization. PeerJ. 2023;11:e16359.
136. Geng Y, Long X, Zhang Y, et al. FTO-targeted siRNA delivery by MSC-derived exosomes synergistically alleviates dopaminergic neuronal death in Parkinson's disease via m6A-dependent regulation of ATM mRNA. J Transl Med. 2023;21:652.
137. Costa-Ferro ZSM, Cunha RS, Rossi EA, et al. Extracellular vesicles derived from mesenchymal stem cells alleviate depressive-like behavior in a rat model of chronic stress. Life Sci. 2025;366-367:123479.
138. Website ClinicalTrials.gov Home Page. Available from https://clinicaltrials.gov/ [accessed 22 May 2026].
139. Xie X, Song Q, Dai C, et al. Clinical safety and efficacy of allogenic human adipose mesenchymal stromal cells-derived exosomes in patients with mild to moderate Alzheimer’s disease: a phase I/II clinical trial. Gen Psychiatr. 2023;36:e101143.
140. Thakur A, Rai D. Global requirements for manufacturing and validation of clinical grade extracellular vesicles. J Liq Biopsy. 2024;6:100278.
141. Kim EH, Son JP, Oh GS, et al. Clinical scale MSC-derived extracellular vesicles enhance poststroke neuroplasticity in rodents and non-human primates. J Extracell Vesicles. 2025;14:e70110.
142. Claridge B, Lozano J, Poh QH, Greening DW. Development of extracellular vesicle therapeutics: challenges, considerations, and opportunities. Front Cell Dev Biol. 2021;9:734720.
143. Ciullo A, Li C, Li L, et al. Biodistribution of unmodified cardiosphere-derived cell extracellular vesicles using single RNA tracing. J Extracell Vesicles. 2022;11:e12178.
144. Zhang T, Zhang L, Ma X, Song W. The tiny giants of regeneration: MSC-derived extracellular vesicles as next-generation therapeutics. Front Cell Dev Biol. 2025;13:1612589.
145. Wiklander OP, Nordin JZ, O’Loughlin A, et al. Extracellular vesicle in vivo biodistribution is determined by cell source, route of administration and targeting. J Extracell Vesicles. 2015;4:26316.
146. Silva AKA, Morille M, Piffoux M, et al. Development of extracellular vesicle-based medicinal products: a position paper of the group “Extracellular Vesicle translatiOn to clinicaL perspectiVEs - EVOLVE France”. Adv Drug Deliv Rev. 2021;179:114001.
147. de Almeida Fuzeta M, Gonçalves PP, Fernandes-Platzgummer A, Cabral JMS, Bernardes N, da Silva CL. From promise to reality: bioengineering strategies to enhance the therapeutic potential of extracellular vesicles. Bioengineering. 2022;9:675.
148. Reiner AT, Witwer KW, van Balkom BWM, et al. Concise review: developing best-practice models for the therapeutic use of extracellular vesicles. Stem Cells Transl Med. 2017;6:1730-9.
149. Tan F, Li X, Wang Z, Li J, Shahzad K, Zheng J. Clinical applications of stem cell-derived exosomes. Signal Transduct Target Ther. 2024;9:17.
Cite This Article
How to Cite
Download Citation
Export Citation File:
Type of Import
Tips on Downloading Citation
Citation Manager File Format
Type of Import
Direct Import: When the Direct Import option is selected (the default state), a dialogue box will give you the option to Save or Open the downloaded citation data. Choosing Open will either launch your citation manager or give you a choice of applications with which to use the metadata. The Save option saves the file locally for later use.
Indirect Import: When the Indirect Import option is selected, the metadata is displayed and may be copied and pasted as needed.
About This Article
Copyright

Data & Comments
Data
0












Comments
Comments must be written in English. Spam, offensive content, impersonation, and private information will not be permitted. If any comment is reported and identified as inappropriate content by OAE staff, the comment will be removed without notice. If you have any queries or need any help, please contact us at support@oaepublish.com.