


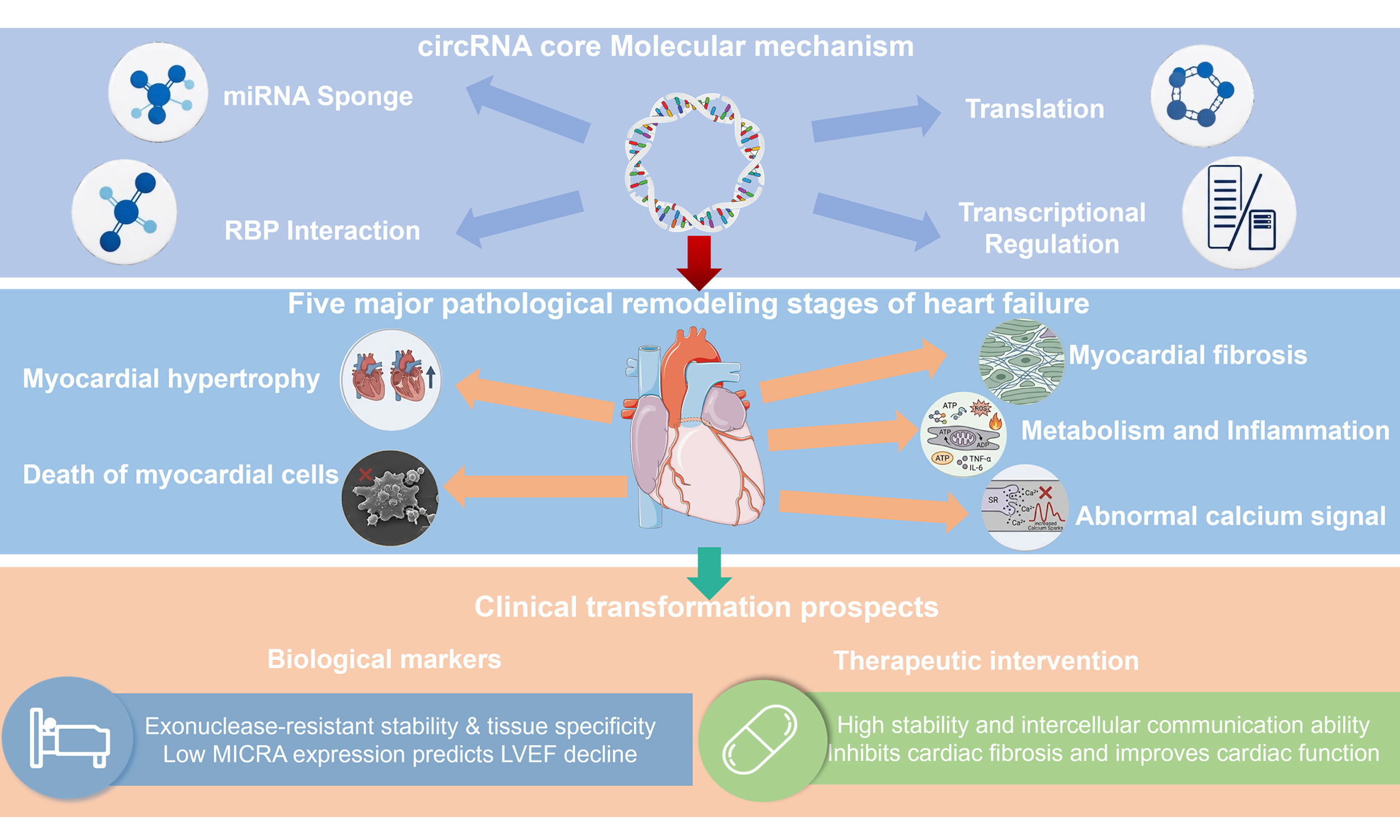
Circular RNAs: key orchestrators of pathological remodeling in heart failure
 0
0 
Abstract
Heart failure (HF) is a complex syndrome driven by structural remodeling, fibrosis, metabolic imbalance, and oxidative stress. Circular RNAs (circRNAs) have emerged as pivotal regulators in HF, distinguished by their covalently closed structure, stability, and tissue specificity. While they modulate myocardial hypertrophy, fibrosis, and metabolic pathways through mechanisms including microRNA (miRNA) sponging, protein interactions, and transcriptional regulation, emerging evidence highlights their critical role in regulating intracellular Ca2+ homeostasis and excitation-contraction (E-C) coupling. By modulating key calcium-handling proteins, circRNAs directly influence cardiomyocyte contractility and arrhythmia susceptibility, linking RNA networks to dynamic cardiac function beyond structural remodeling. Beyond their mechanistic roles, the inherent stability and tissue-specificity of circRNAs position them as highly promising biomarkers for early diagnosis, risk stratification, and therapeutic monitoring. Furthermore, their therapeutic potential is rapidly unfolding; strategies involving extracellular vesicle delivery or targeted silencing offer viable cell-free approaches to inhibit fibrosis and restore cardiac function. This review systematically dissects the multifaceted roles of circRNAs in HF, from their fundamental impact on pathophysiology to their translational journey as biomarkers and therapeutic targets. We also explore current challenges and future directions, providing a comprehensive theoretical framework for the development of circRNA-based precision interventions for heart failure.
Keywords
INTRODUCTION
Cardiovascular diseases, particularly Heart failure (HF), have become a major global public health burden. Recent data indicate that HF affects more than 55 million individuals worldwide[1], with 5-year survival rates varying across clinical phenotypes and regions - generally ranging from approximately 40% to 60%[2] - yet consistently remaining poor and comparable to, or even worse than, several major malignancies, underscoring the substantial mortality burden associated with the disease. The progression to HF is driven by adverse cardiac remodeling, which ultimately causes a decline in cardiac output[3] and functional deterioration. Consequently, although existing biomarkers such as B-type natriuretic peptide (BNP) are widely used for diagnosis and risk stratification, the complex pathophysiology of HF underscores the pressing need for more sensitive multidimensional molecular biomarkers for early screening, disease monitoring, and targeted therapy.
Non-coding RNAs (ncRNAs) play critical regulatory roles in cardiovascular development, homeostasis, and disease progression[4]. Circular RNAs (circRNAs) are a rapidly emerging class of ncRNAs characterized by a covalently closed loop structure lacking both a 5’cap and a 3’poly(A) tail, conferring high stability. Early studies considered circRNAs as by-products of splicing errors[5]; however, since the significant role of circRNA CDR1 antisense (CDR1as) in neural development was reported, research on circRNAs has accelerated[6]. With the development of RNA-seq technology[7] and specific bioinformatics algorithms, over 30,000 circRNAs have been identified across multiple species[8], some of which are highly conserved in humans, suggesting potential functional importance. Several bioinformatics resources have been developed to systematically annotate circRNAs, providing valuable tools for functional studies and potential clinical applications. These databases serve complementary purposes: the Chinese Science Citation Database (CSCD) enables visualization of circRNA structures and back-splicing junctions, circNet integrates circRNA-microRNA (miRNA)-messenger RNA (mRNA) regulatory networks, Circ2Disease focuses on experimentally validated disease associations, and exoRBase profiles circRNAs in human biofluids, facilitating non-invasive biomarker discovery. It is important to note that these resources differ in scope and function, and their accessibility and active maintenance should be verified, as database availability may change over time. Specifically, the databases mentioned above were carefully verified and are currently active and accessible online[9].
Although emerging studies have revealed the involvement of circRNAs in cardiac diseases, comprehensive reviews specifically focused on HF remain limited. Importantly, circRNAs not only modulate structural remodeling processes such as myocardial hypertrophy, fibrosis, and metabolic reprogramming, but also regulate intracellular Ca2+ homeostasis and excitation-contraction (E-C) coupling, linking RNA regulatory networks to cardiac contractility and arrhythmogenic susceptibility. This review will summarize the biogenesis, molecular functions, and mechanisms of circRNAs, with particular emphasis on their regulatory roles in myocardial hypertrophy, cardiomyocyte death, myocardial proliferation and regeneration, myocardial fibrosis, inflammation and metabolic remodeling, and calcium signaling pathways. Furthermore, we discuss their potential clinical applications, analyze current opportunities and limitations, and provide a reference for future translational research of circRNAs in HF.
BIOSYNTHESIS AND MOLECULAR FUNCTION OF CIRCULAR RNA
In this section, we provide an overview of the biosynthesis and molecular functions of circRNAs, including their formation mechanisms and major functional modes. The overall structural framework and key regulatory features are illustrated in Figure 1, and the specific mechanisms will be discussed in detail in the following subsections.
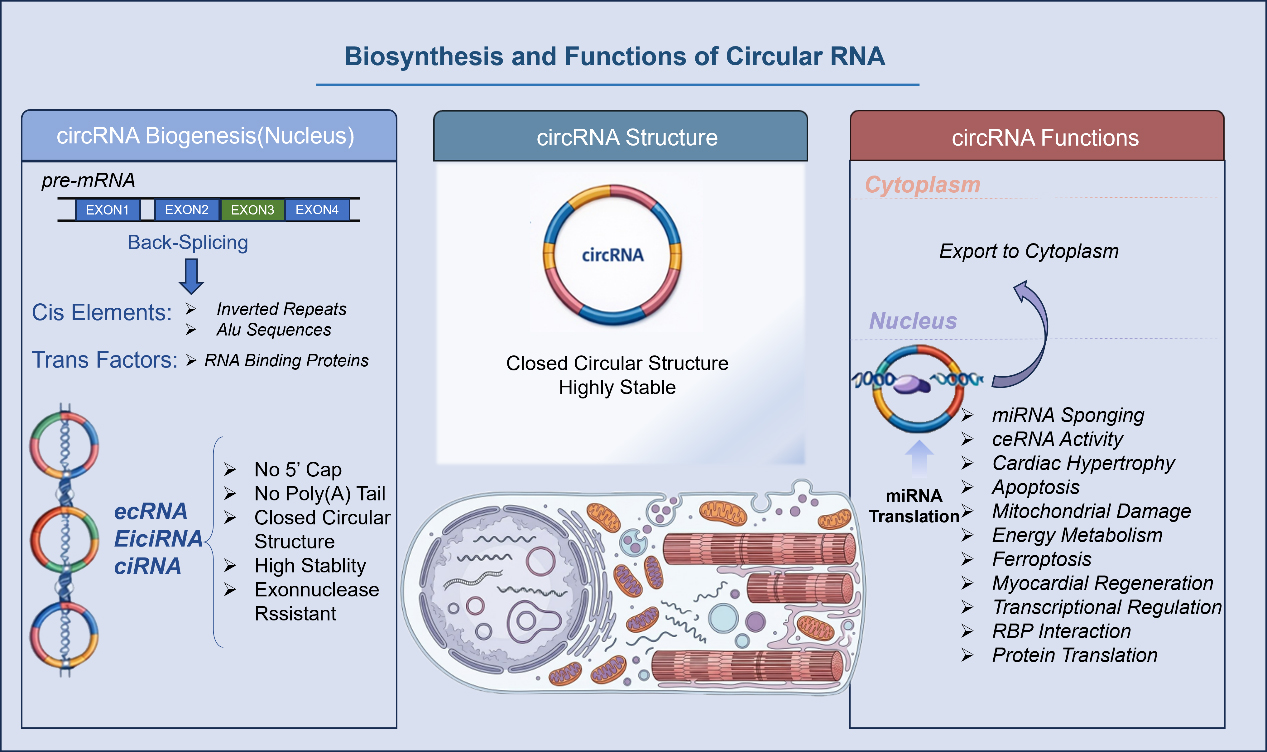
Figure 1. Biosynthesis and Functions of Circular RNA. miRNA: MicroRNA; ceRNA: competing endogenous RNA; circRNA: circular RNA.
Formation mechanism and classification
CircRNAs are endogenous single-stranded RNAs formed from precursor mRNAs through a non-canonical process known as back-splicing, in which a downstream 5’splice site is covalently linked to an upstream 3’splice site to generate a stable, closed circular structure[10,11]. Lacking both a 5’cap and a 3’poly(A) tail, this covalent loop renders circRNAs highly resistant to exonuclease-mediated degradation, giving them stability that far exceeds their linear RNA counterparts[12]. The biogenesis of circRNAs is facilitated by two primary drivers: cis-acting elements, such as inverted repeat sequences (e.g., Alu repeats) within flanking introns that bring splice sites into proximity, and trans-acting RNA-binding proteins (RBPs) that “bridge” distal exons to promote circularization.
This process yields a diverse array of circRNA species. The most common are exonic circRNAs (ecRNAs), composed entirely of one or more exons. Other forms include exon-intron circRNAs (EIciRNAs), which retain intronic sequence, and circular intronic RNAs (ciRNAs), derived solely from introns. Functionally, circRNAs exhibit pronounced tissue-specific and developmental stage-specific expression patterns. While generated in the nucleus, many are exported to the cytoplasm to execute their functions. There, their unique structure enables multi-layered interactions with miRNAs, RBPs, and even DNA, establishing them as critical components of gene regulatory networks[11].
Main molecular functions
miRNA sponge effect
The miRNA sponge effect is a primary mechanism by which circRNAs exert post-transcriptional control[13]. Through specific binding sites, circRNAs competitively sequester miRNAs, reducing their availability to target mRNAs and thereby de-repressing gene expression - a concept known as the competing endogenous RNA (ceRNA) relationship[14,15].
This regulatory axis is a recurring motif across a spectrum of cardiac pathologies. In myocardial hypertrophy, for instance, axes such as cardiac hypertrophy-related circRNA (circRNA CHRC)[16], circCacna1c[17], and circ_Larp4b[18] directly contribute to the hypertrophic response. Similarly, circRNAs critically modulate cardiomyocyte apoptosis and injury responses, where molecules {e.g., cerebellar degeneration-related protein 1 gene (CDR1) and CDR1 antisense RNA (CDR1as)[19]and circSnap47 [a circular RNA derived from the SNAP47 (synaptosome associated protein 47) gene][20]} regulate cell fate through distinct miRNA-dependent pathways, and some can form complexes with multiple miRNAs[21]. The sponge mechanism also extends to metabolic remodeling {e.g., circ-SNRK [a circular RNA derived from the SNRK (SNF-related serine/threonine kinase) gene][22]}, ferroptosis[23] {e.g., circSnx12 [a circular RNA derived from the SNX12 (sorting nexin 12) gene]}, and the function of non-myocyte cells (e.g., cardiac endothelial cells)[24]. In the context of myocardial regeneration, circNfix [a circular RNA derived from the Nfix (nuclear factor I X) gene][25] exemplifies a dual-function model by not only sponging miR-214 but also interacting with RBPs. High-throughput studies further reveal that these interactions form complex networks, with hub circRNAs, such as those from the titin (TTN) gene[26], potentially regulating numerous miRNAs simultaneously.
Overall, circRNAs, through the miRNA-sponge effect, form a complex regulatory network spanning multiple pathological processes, including myocardial hypertrophy, apoptosis, mitochondrial damage, energy metabolism disorders, ferroptosis, and myocardial regeneration. This represents an important direction for understanding the mechanisms of cardiovascular disease and developing precision treatment strategies. Together, these findings highlight the canonical circRNA-miRNA-mRNA regulatory axis as a central pathway in cardiovascular disease, with numerous representative examples summarized in Table 1[27-43].
circRNA-miRNA-mRNA regulatory axes involved in cardiac remodeling
| circRNA | Host gene | miRNA sponged | Target genes/pathways | Pathological process | Expression pattern | Experimental model | Reference |
| circ-SNRK | SNRK | miR-33a | SNRK → AMPK signaling | Inflammation & metabolic regulation | Downregulated | Ischemia/metabolic stress | [22] |
| circSlc8a1 | SLC8A1 (NCX1) | miR-133a | SRF, CTGF, ADRB1 | Cardiac hypertrophy | Upregulated | TAC; Ang II | [27] |
| circDdx60 | Ddx60 | miR-195 | CDK1, Cyclin D1 | Cardiac hypertrophy | Upregulated | Ang II; cardiomyocytes | [28] |
| circ-TLR4 | TLR4 | miR-23a | PPARα | Cardiac hypertrophy | Upregulated | TAC model | [29] |
| circ_0040414 | - | miR-199b-5p | CDK9 | Cardiomyocyte apoptosis | Upregulated | Hypoxia-induced apoptosis | [30] |
| circ_0030042 | - | miR-542-3p | IGF1R | Cardiomyocyte apoptosis | Upregulated | Hypoxia/OGD | [31] |
| circ_006332 | - | miR-1265 | Bcl-2/apoptotic pathways | Cardiomyocyte apoptosis | Upregulated | OGD model | [32] |
| CNEACR (mmu_circ_000338) | - | miR-103-3p | PGAM5 | Cardiomyocyte necrosis | Upregulated | Mitochondrial fragmentation model | [33] |
| circHipk3 | HIPK3 | miR-133a | CTGF, SIRT1 | Cardiomyocyte proliferation | Upregulated | MI regeneration models | [34] |
| circPVT1 | PVT1 | miR-125b | ET-1, pro-fibrotic genes | Cardiac fibrosis | Upregulated | Cardiac fibroblasts | [35] |
| circRNA_000203 | MYH7 | miR-26b-5p; miR-140-3p | CTGF, Col1a2 | Fibrosis | Upregulated | TAC; fibroblasts | [15] |
| circ_0036176 | - | miR-218-5p | Interleukin/ECM pathways | Fibrosis | Upregulated | Cardiac fibroblasts | [36] |
| circNSD1 (circ-0000465) | NSD1 | miR-143-3p | SULF1 → Wnt/β-catenin | Fibrosis | Upregulated | Fibroblast activation model | [37] |
| circNFIB | NFIB | miR-433 | AZIN1 | Anti-fibrotic | Downregulated | Fibrosis models | [38] |
| circYap | YAP | miR-21-3p | TGF-β/Smad signaling | Anti-fibrotic | Downregulated | Cardiac fibroblasts | [39] |
| circHelz | HELZ | miR-133a | TGF-β1, Col1a1 | Fibrosis | Upregulated | Fibroblasts | [40] |
| circ_0002295 | - | miR-29b | Col1/Col3 pathway | Fibrosis | Upregulated | Fibroblasts | [41] |
| circSamd4 | Samd4 | miR-138-5p | SIRT1 | Metabolic/inflammatory remodeling | Upregulated | Lipotoxic cardiomyocyte model | [42] |
| FEACR | - | miR-432 | PCK1/FA metabolism | Metabolic remodeling | Upregulated | Fatty-acid metabolic stress | [43] |
RNA-binding protein
Beyond miRNA sponging, direct interaction between circRNAs and RBPs represents another pivotal mode of pathological regulation[44]. With over 800 RBPs[27] identified in the human genome, these interactions are integral to the entire circRNA lifecycle, from biogenesis to functional execution.
In the “RBP scaffold” mechanism, circRNAs provide a platform to assemble, stabilize, or regulate protein complexes. For instance, circ-Foxo3 can form a ternary complex with cyclin-dependent kinase 2 (CDK2) and p21 to inhibit cell cycle progression[45], while circNfix facilitates the degradation of the RBP Y-box binding protein 1 (Ybx1) to limit cardiomyocyte proliferation[25]. Conversely, in the “RBP sponge” mechanism, circRNAs sequester RBPs blocking their function, as seen with circCmss1, which binds eukaryotic translation initiation factor 4A3 (EIF4A3) to promote ferroptosis[46]. This functional axis extends to epigenetic regulation, where circRNAs such as autophagy-associated circRNA (ACR) can bind to DNA methyltransferases [e.g., DNA methyltransferase 3B (Dnmt3B)] to modulate gene expression[30]. Furthermore, these interactions are critical in modulating cardiac hypertrophy and stress responses; for example, circ-Ddx60 directly binds eukaryotic elongation factor 2 (eEF2) to enhance AMP-activated protein kinase (AMPK) signaling and exert an anti-hypertrophic effect[28].
Overall, circRNA-RBP interactions exhibit high functional diversity, covering multiple pathological processes including cell cycle regulation, protein degradation, ferroptosis, autophagy, metabolic regulation, and stress adaptation. The circRNA-RBP axis has emerged as a key component in understanding the development of cardiovascular diseases and offers new potential targets for regulating myocardial regeneration, inhibiting pathological hypertrophy, and intervening in myocardial injury. The stable, closed conformation of circRNAs is thought to confer greater protein-binding capacity than linear RNAs, enabling two primary mechanisms of action: acting as protein scaffolds or as protein sponges, with key examples detailed in Table 2[47-51].
circRNA-RBP interactions involved in cardiac remodeling
| circRNA | Host gene | Binding RBP | Mechanistic category | Downstream targets/pathways | Pathological process | Experimental model | Reference |
| circNfix | Nfix | Ybx1/Nedd4l | RBP degradation; ubiquitination regulation | GSK3β-β-catenin; Ybx1 stability | Blocks cardiomyocyte proliferation; limits regeneration | MI model | [25] |
| circRNA_000203 | MYH7 | Transcription factor-promoter binding enhancer | Scaffold-enhancing CTGF promoter activity | CTGF, Col1a2 ↑ | Pro-fibrotic | TAC; cardiac fibroblasts | [36] |
| circNSD1 (circ-0000465) | NSD1 | Chromatin-associated protein pathway | ceRNA + protein-modulating axis | SULF1 → Wnt/β-catenin activation | Pro-fibrotic | Cardiac fibroblasts | [37] |
| circYap | YAP | TEAD/YAP pathway proteins | Scaffold-enhancing YAP-TEAD transcription | YAP-TEAD downstream genes | Anti-fibrotic | Cardiac fibroblast models | [39] |
| ACR | - | AKT | Direct protein activation | Pink1-Parkin pathway | Reduces autophagy-related cell death | Hypoxia model | [47] |
| circSlc8a1 | SLC8A1 (NCX1) | QKI (biogenesis regulator) | RBP-regulated circRNA formation | miR-133a → SRF/CTGF/ADRB1 | Ca2+ handling; hypertrophy | TAC/AAV | [48] |
| circCDYL | CDYL | Translation machinery (ribosome-binding) | Noncanonical peptide translation (tCDYL-100) | REST-CDYL-EHMT2 complex | Promotes hypertrophy | Ang II model | [49] |
| circWhsc1 | Whsc1 | STAT3 | Activation of STAT3 signaling | TRIM59/Cyclin B2 | Promotes cardiac repair; regeneration | MI + extracellular vesicles | [50] |
| circ23679 | - | RBM24 | RBP-mediated circRNA biogenesis | miR-15b-5p axis | Anti-apoptotic | Cardiomyocyte models; mouse hearts | [51] |
Translation potential
While traditionally classified as non-coding, a growing body of evidence reveals that many circRNAs possess translational capacity. This function is driven by cap-independent mechanisms, such as the presence of Internal Ribosome Entry Sites (IRES) or N6-methyladenosine (m6A) modifications[52], which allow ribosomes to initiate translation on a closed-loop template. The discovery of adenine-uracil (AU)-rich IRES-like elements that drive efficient translation has added a disruptive new dimension to circRNA functional research, revealing a novel layer of protein-coding potential independent of linear transcripts.
The functional relevance of these circRNA-encoded peptides is increasingly evident in cardiac pathology. In myocardial fibrosis, for instance, the peptide Myo9a-208, translated from circ_0036176, exhibits anti-fibrotic properties[36], and its production is fascinatingly regulated by a miRNA. In myocardial hypertrophy, the circCDYL [a circular RNA derived from the CDYL (chromodomain Y-like) gene]-encoded peptide tCDYL-100 is thought to competitively modulate chromatin regulation[49]. Pathogenic peptides have also been identified; the Nlgn173 peptide, translated from circNlgn [a circular RNA derived from the Nlgn (neuroligin) gene][53], promotes collagen deposition and cardiac dysfunction, making it a potential therapeutic target[54]. Furthermore, in cardiac energy metabolism, the circSLC8a1-derived peptide SLC8a1-604 impairs mitochondrial function and adenosine triphosphate (ATP) production[45], directly contributing to ventricular dysfunction. These examples, further detailed in Table 3[55-57], highlight how circRNA-derived peptides are key players in diverse pathological processes.
circRNA-encoded peptides involved in cardiac remodeling
| circRNA | Host gene | Encoded peptide | Mechanistic category | Downstream pathways/targets | Pathological role in cardiac remodeling | Experimental model | Reference |
| circCDYL | CDYL | tCDYL-100 peptide | Peptide binds to REST-CDYL-EHMT2 complex; enhances H3K9 methylation | REST/CDYL/EHMT2 epigenetic repression | Promotes pathological cardiac hypertrophy | Ang II-induced hypertrophy models (in vivo & in vitro) | [49] |
| circZNF609 | ZNF609 | ZNF609-derived peptide | Translated via an IRES-like structure; regulates cytoskeletal remodeling | Cytoskeletal and proliferation-related pathways | Regulates muscle cell proliferation; potential influence on structural remodeling | Skeletal & cardiac muscle models | [55] |
| circMAPK1 | MAPK1 | MAPK1-109aa peptide | Competes with MAPK1 translation; fine-tunes ERK/MAPK signaling | ERK/MAPK pathway modulation | Modulates cell survival; may influence ischemic remodeling | Hypoxia / cellular stress models | [56] |
| circSHPRH | SHPRH | SHPRH-146aa peptide | Protects SHPRH protein from degradation; regulates DNA repair | DNA repair and cell-protection pathways | Protects against cellular injury; may affect I/R remodeling | I/R and oxidative stress models | [57] |
In summary, the translational potential of circRNAs offers a paradigm-shifting perspective on myocardial remodeling. With the maturation of ribosome profiling (Ribo-seq), mass spectrometry, and long-read sequencing technologies, the landscape of the circRNA translatome will continue to expand. The encoded micropeptides are emerging as central nodes in disease progression and promising new targets for anti-fibrosis and anti-heart failure therapies.
Transcriptional regulation
Beyond their cytoplasmic roles, a distinct class of nuclear-localized circRNAs has emerged, playing a crucial role in cardiovascular pathology by directly regulating transcription and epigenetics. This functional arm is largely composed of ciRNAs and EIciRNAs, which can enhance the transcription of their own host genes in a cis-acting manner. The core mechanism often involves interacting with the U1 small nuclear ribonucleoprotein (U1 snRNP) and RNA Polymerase II (Pol II) machinery to boost transcriptional activity, establishing circRNAs as direct, gene-specific transcriptional regulators[58].
A prime example in the cardiovascular system is the ACR, which is significantly downregulated in ischemia-reperfusion injury and exerts a potent cardioprotective effect[47]. Crucially, this function stems not from miRNA sponging but from its nuclear epigenetic capacity: ACR directly binds the DNA methyltransferase Dnmt3B, preventing methylation of the Pink1 promoter and thereby sustaining its expression. This was a landmark finding, demonstrating for the first time that a circRNA could epigenetically regulate a key cardioprotective gene, intervening at a far more upstream node of myocardial stress responses.
The regulatory spectrum of nuclear circRNAs continues to expand. More recent evidence reveals their capacity to act as modular scaffolds, promoting the assembly of transcription factors and chromatin remodeling complexes [e.g., SWItch/Sucrose Non-Fermentable (SWI/SNF) complex] to influence key stress-response and fibrotic genes[59,60]. These findings collectively establish that circRNA functions extend to the highest levels of gene regulation - including chromatin state and 3D genome architecture - offering profound implications for understanding the molecular underpinnings of myocardial remodeling.
CIRCRNA IN THE MOLECULAR PATHOLOGICAL MECHANISMS OF HEART FAILURE
With the development of transcriptomic sequencing technology, numerous studies have confirmed that circRNAs have broader and more refined regulatory functions in cardiovascular diseases than previously thought[61]. HF, as a multi-pathogenic final pathway, involves a complex molecular network during its development, including myocardial hypertrophy, cardiomyocyte death, myocardial fibrosis, inflammatory responses, and remodeling of energy metabolism. circRNAs exhibit high stability, tissue specificity and dynamic expression patterns, and they are involved in almost all critical pathological processes[62,63]. Therefore, they not only play a crucial role as an important node in the molecular regulatory network of HF, but are also gradually emerging as potential diagnostic markers and therapeutic targets.
Regulation of cardiomyocyte hypertrophy
Persistent pathological myocardial hypertrophy is a key early event in the development of HF and a primary therapeutic target. While initially an adaptive response, sustained stress drives the heart from compensation to decompensation. Numerous studies[9] have now confirmed the crucial role of circRNAs within the multi-layered regulatory network of hypertrophy, providing new directions for understanding myocardial remodeling.
Circular RNAs act as miRNA sponges in cardiomyocyte hypertrophy
The miRNA sponge or ceRNA mechanism is a cornerstone of circRNA function in hypertrophy. This principle was established in early studies of molecules such as heart-related circRNA (HRCR), which was found to regulate the miR-223/ARC axis[64]. Since then, research has rapidly expanded this network, identifying a multitude of circRNAs - such as circRNA CHRC[5,16], circSlc8a1[11], circCacna1c[17], and circ_Larp4b[18] - that sponge distinct miRNAs to modulate key pro-hypertrophic transcription factors [e.g., Krüppel-like factor 15 (KLF15), Myocyte enhancer factor 2C (Mef2c)] and signaling pathways. This positions the circRNA-miRNA axis as a pivotal and highly plastic regulatory hub in pathological remodeling.
Interaction of circular RNA with RBP in myocardial hypertrophy
Beyond sponging miRNAs, circRNAs directly engage with RBPs to regulate hypertrophy, often through two distinct modes. Some, including circ-Ddx60[28], act as protein-binding partners, directly enhancing protective signaling pathways such as AMPK-eEF2 signaling (adenosine monophosphate-activated protein kinase-eEF2) in response to physiological stress. Others, such as circ-Toll-like receptor 4 (TLR4)[29], function as molecular scaffolds, recruiting proteins [e.g., Fused in sarcoma (FUS)] to increase the stability and expression of pro-hypertrophy inflammatory genes (e.g., TLR4). These findings reveal an important layer of regulation in which circRNAs influence hypertrophy by modulating protein stability and directly signaling.
Circular RNA encoding protein in myocardial hypertrophy
A paradigm-shifting mechanism involves the translation of circRNAs into functional micropeptides. The study of circCDYL[49] provides the quintessential example in hypertrophy. This circRNA, upregulated by hypertrophic stimuli, is translated into a small peptide (tCDYL-100) that is itself sufficient to drive a pro-hypertrophic phenotype. Mechanistically, tCDYL-100 achieves this by disrupting the REST-CDYL (Chromodomain Y-Like)-Euchromatic histone lysine methyltransferase 2 (EHMT2) epigenetic repressor complex, thereby de-repressing key pro-hypertrophy genes. This landmark discovery not only confirms the coding potential of circRNAs but also directly links it to epigenetic dysregulation in cardiac remodeling, opening an entirely new field of inquiry.
Regulation of cardiomyocyte death
Cardiomyocyte death, encompassing apoptosis, necroptosis, ferroptosis, and dysregulated autophagy, is a core driver of adverse remodeling and the progression to HF. Recent studies reveal that circRNAs are deeply integrated into these cell death pathways, employing a diverse toolkit of molecular mechanisms that extend far beyond miRNA sponging to include RBP interactions, epigenetic control, and the maintenance of mitochondrial homeostasis[65].
In the regulation of apoptosis, the miRNA sponge mechanism remains a prominent theme. circRNAs such as circ_0040414[30], hsa_circ_0097435[21], and circ23679[51] have all been shown to modulate canonical life/death signaling pathways [e.g., phosphatase and tensin homolog (PTEN)/protein kinase B (AKT) signaling pathway, mitogen-activated protein kinase (MAPK) signaling] by sequestering specific miRNAs under various stress conditions[66]. This mechanism also extends to inflammatory cell death such as pyroptosis, where specific circRNAs have been shown to exacerbate myocardial damage by modulating inflammatory pathways[31,32]. In necroptosis, however, protein interactions take center stage. For instance, cardiac- necroptosis-associated circRNA (CNEACR)[33] directly binds to histone deacetylase 7 (HDAC7), preventing its inhibition of a key transcription factor and thereby blocking the RIPK3-mediated (receptor-interacting serine/threonine-protein kinase 3-mediated) necroptotic pathway, showcasing a precise protein-localization-based control mechanism.
circRNAs also play a crucial role in regulating metabolic forms of cell death. In ferroptosis, circRNAs such as ferroptosis‐associated circRNA (FEACR)[43] can act as metabolic regulators, binding to and stabilizing key enzymes to maintain redox balance and prevent iron-dependent cell death. In the context of autophagy and mitochondrial homeostasis, circRNAs demonstrate remarkable versatility. Some, including the nuclear circRNA ACR[47], epigenetically regulate the transcription of key mitochondrial protective genes. Others, such as circSamd4 [a circular RNA derived from the SAMD4 (sterile alpha motif domain containing 4) gene][28], function in the cytoplasm to directly maintain mitochondrial protein localization and function, thereby preventing mitochondrial instability and cell death.
In summary, circRNAs orchestrate the complex landscape of cardiomyocyte death through a multi-dimensional toolkit, including miRNA sponging, RBP scaffolding, and epigenetic modulation. They are closely linked to classical signaling pathways such as PTEN/AKT, RIPK3, and the NAMPT/Sirt1 axis (nicotinamide phosphoribosyltransferase/sirtuin 1), underscoring their central role in cardiac injury and repair. Further research, particularly in large animal models, is needed to translate these mechanistic insights into precise, circRNA-based therapeutic interventions for HF.
The role of CircRNA in myocardial proliferation and regeneration
Enhancing the extremely limited regenerative capacity of the adult heart is a core goal of HF research. While strategies such as cell transplantation exist, their clinical efficacy is limited. Consequently, identifying novel molecular targets to boost endogenous regeneration is a critical research direction. Against this backdrop, circRNAs have emerged as key regulatory molecules in cardiac proliferation and repair.
The regulatory landscape of circRNAs in regeneration is characterized by a fascinating duality, with distinct species acting as either potent inhibitors or promoters of cardiomyocyte proliferation. circNfix[25] is the quintessential “inhibitory” circRNA. It is highly expressed in the adult heart and restricts proliferation via a dual mechanism: it facilitates the ubiquitination and degradation of key cell-cycle proteins, and it sponges miR-214 to inhibit pro-regenerative β-catenin signaling. Consequently, inhibiting circNfix has been shown to enhance cardiomyocyte proliferation and improve cardiac function post-myocardial infarction (post-MI), highlighting its significant therapeutic potential. Conversely, circHipk3[25,67] is a classic “pro-regenerative” circRNA, highly expressed during early cardiac development. It promotes proliferation and angiogenesis[68] through a multi-pronged approach, including stabilizing the neurogenic locus notch homolog protein 1 (Notch1) signaling pathway and sponging miR-133a to de-repress downstream growth factors. More recently, circCDYL has also been identified as a pro-proliferative molecule that functions primarily by sponging miR-4793-5p to promote myocardial repair[69].
In summary, molecules such as circNfix, circHipk3, and circCDYL represent distinct but archetypal modes of circRNA action in cardiac regeneration, from dual-mechanism inhibition to multi-target activation. While this field is still nascent, these studies reveal the crucial role of circRNAs in governing the heart's regenerative potential. They provide a strong theoretical and practical foundation for the future development of RNA-targeted therapies aimed at repairing the injured myocardium and ultimately treating HF.
circRNA and myocardial fibrosis
Myocardial fibrosis, characterized by excessive extracellular matrix deposition, is a core pathological driver of cardiac stiffness and the progression to HF. In this complex process, circRNAs have been shown to play a key role, modulating fibrosis through multiple signaling pathways[70].
circRNAs regulate fibrosis by targeting a web of crucial signaling cascades. Within the pivotal transforming growth factor-beta (TGF-β) signaling pathway, for instance, some circRNAs such as circPVT1 [a circular RNA originating from back-splicing of the PVT1 (plasmacytoma variant translocation 1) gene][35] are pro-fibrotic, while others such as circSMAD3 [circSMAD3, a circular RNA derived from the SMAD3 (SMAD family member 3) gene][71] can be inhibitory. This pathway is also targeted by molecules such as circSMAD4[72]. The complexity is further underscored by different circRNAs from the same gene locus exhibiting opposing functions; for instance, the myosin IXA (MYO9A) gene produces both pro- and anti-fibrotic circRNAs[36]. Beyond TGF-β, circRNAs also target other key cascades, such as the Wnt/β-catenin pathway, which is activated by circNSD1[37], and inflammatory feedback loops driven by peptides such as Nlgn173[53]. The regulatory landscape is further enriched by numerous other circRNAs that modulate fibrosis by targeting distinct molecular nodes. These include those that modulate the crucial Yes-associated protein 1 (YAP) signaling pathway (e.g., circYap[39] and circHelz[40]), and others that act through different axes to exert either anti-fibrotic (e.g., circNFIB[38], circ-sh3rf3[73]) or pro-fibrotic (e.g., circ_0002295[41]) effects.
In summary, circRNAs act as key upstream regulators in myocardial fibrosis by modulating a dense network of signaling pathways including TGF-β/Smad, Wnt/β-catenin, and YAP. Crucially, these studies reveal the pronounced bidirectional nature of circRNA regulation. From pro-fibrotic players (e.g., circPVT1 and circHelz) to anti-fibrotic molecules (e.g., circSMAD3 and circYap), this duality is a recurring theme. This provides a strong foundation for developing highly specific, RNA-targeted therapeutic strategies that either inhibit pro-fibrotic circRNAs or augment protective circRNAs to combat HF.
The role of CircRNA in inflammation and metabolic remodeling
Beyond structural changes, HF is driven by a complex interplay of metabolic imbalance, mitochondrial dysfunction, oxidative stress, ferroptosis, and inflammation. Recent studies have positioned circRNAs as crucial molecular hubs that connect these metabolic and stress pathways to myocardial pathology, employing a range of mechanisms from miRNA sponging to direct protein interaction.
A key theme emerging is the role of specific circRNAs as guardians of cellular homeostasis, particularly against metabolic and oxidative stress. circSamd4[42], for example, is a mitochondrial-localized circRNA that protects cardiomyocytes by reducing mitochondrial reactive oxygen species (ROS), inhibiting apoptosis, and promoting repair. Its mechanism highlights a direct link between circRNA function and the mitigation of oxidative damage. Similarly, FEACR[43] acts as a crucial regulator of ferroptosis, a form of iron-dependent cell death critical in ischemia/reperfusion injury. It achieves this by stabilizing a key enzyme (NAMPT) to ultimately upregulate the ferroptosis inhibitor ferritin heavy chain 1 (FTH1), representing a novel protective axis against ischemia/reperfusion injury (I/R injury). Furthermore, circRNAs such as circ-SNRK[74] contribute to energy metabolism by modulating their host gene’s expression via a miRNA sponge mechanism, thereby protecting myocardial energy balance. These studies collectively demonstrate how circRNAs can maintain metabolic homeostasis and protect the heart from various forms of stress-induced cell death.
In summary, circRNAs are deeply integrated into the networks governing myocardial metabolic homeostasis, mitochondrial function, oxidative stress, and ferroptosis. They can act as potent protective factors, as exemplified by circSamd4, FEACR, and circ-SNRK, helping the myocardium resist injury. However, their dysregulation can also exacerbate energy depletion and cell death, promoting cardiac dysfunction. Incorporating circRNAs into the broader framework of cardiac inflammation and metabolic remodeling will be essential for fully understanding disease progression and developing novel therapeutic interventions for HF.
The role of CircRNA in the calcium signaling pathway
The regulation of calcium (Ca2+) homeostasis is fundamental to cardiac function, and its dysregulation is a hallmark of HF. Recent research has unveiled a new frontier: the direct role of circRNAs in modulating Ca2+ handling, offering a novel perspective on the molecular drivers of HF.
A key mechanism involves circRNAs that are derived from, and subsequently regulate, major Ca2+ handling genes. circRYR2[75], for instance, is a circRNA derived from the ryanodine receptor 2 (RYR2) locus that is downregulated in pathological hypertrophy. Its deficiency perturbs the expression of key proteins such as sarcoplasmic/endoplasmic reticulum Ca2+-ATPase 2a (SERCA2a) and phospholamban (PLN), impairing Ca2+ reuptake and prolonging Ca2+ transients. Conversely, restoring circRYR2 expression improves Ca2+ handling and cardiac function, establishing a direct link between a circRNA and myocardial contractility. Similarly, circSlc8a1[76], derived from the SLC8A1/NCX1 (solute carrier family 8 member A1/sodium-calcium exchanger 1) locus, is crucial for maintaining Ca2+ efflux. Its specific silencing leads to severe HF, while its forced expression is protective. Mechanistically, circSlc8a1 deficiency disrupts NCX1-mediated Ca2+ efflux, triggering diastolic Ca2+ overload and adverse metabolic remodeling, highlighting its essential role in preventing HF progression.
Beyond direct modulation of transport proteins, circRNAs also indirectly influence Ca2+ dynamics by targeting upstream signaling enzymes and pathways. For example, some circRNAs can alter cyclic adenosine monophosphate (cAMP) signaling and PKA/PKC (protein kinase A / protein kinase C) kinase activity by sponging miRNAs that target key enzymes such as adenylate cyclase[34]. Furthermore, in the context of ischemia/reperfusion injury, circRNAs have been shown to modulate neuropeptide signaling[77], which in turn alters the phosphorylation state of Ca2+ handling proteins (e.g., RYR2 and PLN), ultimately impacting cardiomyocyte survival.
In summary, circRNAs form a hierarchical, deeply integrated regulatory network governing myocardial Ca2+ homeostasis. By acting on channels, transporters, and signaling enzymes, they fine-tune Ca2+ dynamics and electromechanical coupling. This emerging perspective does not merely supplement our understanding of hypertrophy and fibrosis; it establishes the circRNA-Ca2+ axis as a new, fundamental pillar of cardiac regulation. This provides a solid theoretical basis for developing novel therapeutic strategies aimed at restoring Ca2+ homeostasis by targeting specific circRNA.
As a final summary, this article systematically reviews the roles of circRNAs in myocardial remodeling and HF. The reported key circRNAs, along with their host genes, mechanisms of action, downstream signaling pathways, and associated pathological processes, are comprehensively summarized in Table 4[78,79]. Collectively, these findings provide a panoramic overview of the multi-level regulatory functions of circRNAs in myocardial hypertrophy, cell death, fibrosis, metabolic remodeling, calcium homeostasis, and myocardial regeneration, highlighting their significance in the pathogenesis and potential therapeutic targeting of cardiovascular disease.
Representative circRNAs involved in cardiac remodeling and heart failure (final unified 7-column format)
| circRNA | Host gene | Pathological process | Mechanistic category | Key downstream targets/pathways | Expression pattern | Experimental model | Reference |
| HRCR | SLC8A1 | Anti-hypertrophy | miRNA sponge | miR-223 → ARC ↑ | Down | TAC | [64] |
| circRNA-CHRC | - | Anti-hypertrophy | miRNA sponge | miR-431-5p → KLF15 ↑ | Down | Ang II hypertrophy | [16] |
| circCacna1c | Cacna1c | Promotes hypertrophy | miRNA sponge | miR-29b-2-5p → NFATc1 ↑ | Up | ISO-induced hypertrophy | [17] |
| circ_Larp4b | Larp4b | Promotes hypertrophy | miRNA sponge | miR-298-5p → MEF2C ↑ | Up | Ang II model | [18] |
| circ-TLR4 | TLR4 | Pro-hypertrophy | miRNA sponge | miR-23a/NF-κB-related pathways | Up | Ang II/LPS | [32] |
| circ-Ddx60 | Ddx60 | Pro-hypertrophy | ceRNA regulatory axis | Hypertrophic gene program activation | Up | TAC/Ang II | [28] |
| circ23679 | - | Anti-apoptosis | miRNA sponge | miR-15b-5p | Up | Cell + mouse models | [51] |
| mmu_circ_000338 (CNEACR) | - | Anti-apoptosis reduces I/R injury | miRNA sponge | miR-545-3p → anti-apoptotic genes | Up | I/R | [33] |
| MFACR | - | Mitochondrial fission-induced apoptosis | miRNA sponge | miR-652-3p → MTP18 ↑ | Up | Ischemia/reperfusion model | [78] |
| circ_006332 | - | Regulates oxidative-stress apoptosis | miRNA sponge | Nrf2-related genes | Up | Oxidative stress model | [32] |
| circ_0040414 | - | Promotes apoptosis under stress | miRNA sponge | Mitochondrial apoptotic regulators | Up | Hypoxia/Ang II | [30] |
| ACR | - | Suppresses autophagy-related cell death | Direct AKT activation/miRNA sponge | Pink1/Parkin pathway | Up | Hypoxia | [47] |
| hsa_circ_0097435 | - | HF-associated apoptosis | AGO2-mediated multi-miRNA binding | Multi-target apoptosis pathways | Up | Human peripheral blood (HF) | [21] |
| circNfix | Nfix | Suppresses proliferation/regeneration | miRNA sponge + RBP degradation | miR-214; Ybx1-Nedd4l axis | High in the adult heart | MI model; in vitro CMs | [25] |
| circHipk3 | Hipk3 | Promotes cardiomyocyte proliferation | miRNA sponge | miR-133a; Akt-Notch pathway | Up | Neonatal heart/CM culture | [68] |
| circCDYL | CDYL | Pro-hypertrophy; epigenetic regulation | Encodes peptide (tCDYL-100) | REST-CDYL-EHMT2 | Up | In vivo and in vitro hypertrophy | [49] |
| circNFIB | NFIB | Anti-fibrotic | miRNA sponge | miR-433 | Down | MI/TGF-β | [38] |
| circ_0036176 | MYO9A | Anti-fibrotic effects | EV-associated regulation | Suppresses fibroblast activation | Upregulated | MI + EV treatment | [36] |
| circYap | YAP | Anti-fibrotic | Protein interaction | TPM4/ACTG binding → inhibition of myofibroblast transition | Down | Pressure overload models | [39] |
| circPVT1 | PVT1 | Promotes fibrosis | miRNA sponge | miR-30a-5p; miR-125b-5p | Up | HF myocardium | [35] |
| circ_0002295 | - | Promotes fibrosis | miRNA sponge | miR-1287 → CXCR2 ↑ | Up | Human MF samples | [41] |
| circHelz | Helz | Promotes fibrosis | ceRNA axis | Collagen synthesis-related genes | Up | CFs | [40] |
| circSNRK | SNRK | Inflammation and metabolic remodeling | miRNA sponge | miR-33 → SNRK ↑ | Down | MI model | [22] |
| circSamd4 | Samd4 | Inflammation and metabolic remodeling | Mitochondrial function regulation | Nrf2-dependent expression; ↓ mitochondrial ROS; ↓ oxidative DNA damage | Highly expressed | MI model; embryonic and neonatal cardiomyocytes | [42] |
| FEACR | - | Inflammation and metabolic remodeling | Ferroptosis regulation via protein interaction | NAMPT stabilization → Sirt1 activation → FOXO1 deacetylation → FTH1 upregulation; | Downregulated | MI model | [43] |
| circSlc8a1 | SLC8A1 (NCX1) | Calcium signaling dysregulation | miRNA sponge | miR-133a → SRF/ADRB1/CTGF | Dynamic | TAC; AAV overexpression/inhibition | [48] |
| circHipk3 | Hipk3 | Calcium signaling modulation | miRNA sponge | miR-17 family → ADCY6 → cAMP/PKA/PKC pathway → Ca2+ handling modulation | Context-dependent | Cardiac remodeling models | [34] |
| mmu_circ_0000021 | - | Calcium signaling in I/R injury | miRNA sponge | miR-143-3p → NPY ↑ → p-RYR2 ↑ | Upregulated | I/R mouse model | [77] |
| circRYR2 | RYR2 | Calcium signaling dysregulation | Ca2+ handling modulation | SERCA2a/PLN/CSQ | Down | TAC; mouse + human samples | [75] |
| MICRA | - | Biomarker predicting LV remodeling after MI | Circulating RNA marker | - | Low → Poor LVEF recovery | Human AMI cohort (n = 472) | [79] |
CLINICAL APPLICATION POTENTIAL OF CIRCRNA
Potential as a Biomarker for Circulation
An ideal biomarker should be a measurable molecule that reflects disease outcome, course, or treatment response. In HF, classic markers such as BNP and N-terminal pro–B-type natriuretic peptide (NT-proBNP) are widely used for this purpose. circRNAs, however, present a next-generation opportunity. Their covalently closed structure confers exceptional resistance to exonuclease degradation, resulting in stability in blood and other bodily fluids that far surpasses linear RNAs[80,81]. This inherent stability, combined with their tissue-specific expression, makes them naturally suited to be robust clinical biomarkers.
Growing evidence now substantiates the clinical value of circRNAs in the diagnosis and risk prediction of HF. Multiple studies report that specific circRNAs are abnormally expressed in the peripheral blood of HF patients[82]. For instance, circ_0040414 is significantly elevated in patients with chronic heart failure (CHF) and participates in pathological signaling via the ceRNA mechanism[83]. Among the circRNAs investigated as prognostic biomarkers, MICRA (myocardial infarction-associated circRNA) currently represents the most prominent example approaching clinical applicability. A prospective cohort study of acute myocardial infarction (AMI) patients demonstrated that low MICRA expression was a significant and independent predictor of subsequent left ventricular ejection fraction (LVEF) decline. To date, no other circRNA candidates have achieved comparable clinical validation or been evaluated in large patient cohorts, highlighting MICRA’s unique role in bridging basic circRNA research and patient-centered prognostic assessment.
With the advancement of high-throughput sequencing, the capacity to identify and quantify specific circulating circRNAs continues to improve. This opens the door for their use in non-invasive diagnosis, disease subtyping, risk prediction, and efficacy monitoring, laying the foundation for a precision medicine approach to HF.
Potential as a therapeutic target
The therapeutic potential of circRNAs is rapidly gaining attention. Their high stability and ability to be packaged into extracellular vesicles (EVs) for intercellular communication[84] provide a feasible pathway for “cell-free” therapeutic strategies[85]. circRNAs released from cardiac cells can be delivered via EVs to target cells, where they can modulate key pathological pathways, including proliferation, apoptosis, and fibrosis[86]. This mechanism offers several advantages for therapeutic delivery, including natural nuclease resistance and the intrinsic targeting capabilities of EVs.
Functional validation for this concept already exists. For instance, EV-delivered circRNAs have been shown in preclinical models to inhibit cardiac fibrosis (e.g., circ_0036176)[36] or to promote cardiomyocyte proliferation and improve cardiac function post-MI (e.g., circWhsc1)[50]. Beyond EV delivery, synthetic circRNA mimetics or antisense oligonucleotides (ASOs) designed to silence pathogenic circRNAs represent another promising therapeutic avenue, offering the potential to reverse pathological remodeling at the molecular level.
However, several technical limitations currently constrain the clinical translation of EV-based circRNA therapies. EV isolation and purification lack universally standardized protocols, and methodological variability can significantly affect vesicle composition and yield. In addition, in vivo targeting efficiency remains suboptimal, as systemically administered EVs are often rapidly cleared and accumulate off-target in organs such as the liver and spleen. Furthermore, large-scale production under good manufacturing practice (GMP) conditions, together with batch-to-batch consistency and rigorous quality control, presents substantial logistical and regulatory challenges. Addressing these issues will be essential to ensure the reproducibility, safety, and scalability of EV-mediated circRNA delivery.
In summary, circRNAs can regulate cardiovascular pathology at multiple levels. Their stability and druggability, particularly via EV-based strategies, provide actionable pathways from molecular mechanism to clinical intervention. By optimizing delivery and ensuring long-term safety, circRNA-based therapies are poised to become a next-generation tool for treating cardiovascular diseases.
Challenges and prospects
Despite their immense potential, the clinical translation of circRNAs faces several key challenges. First, detection and analysis methods lack full standardization, with differences in sample preparation[87], algorithms[88], and quantification strategies hindering cross-study comparisons. Second, the tissue-specific and dynamic expression patterns of circRNAs are not yet fully mapped, making it difficult to define optimal detection and treatment windows[89].
Furthermore, the majority of current mechanistic studies rely on rodent models, whose physiological differences from humans create uncertainty for direct clinical translation. Future work must therefore extend to large animal models and human tissues. Simultaneously, the safety, specificity, and immunogenicity of any circRNA-based therapeutic, whether delivered via EVs[90] or as synthetic molecules, require rigorous preclinical evaluation.
Looking ahead, the combination of high-throughput sequencing, single-cell analysis, and spatial transcriptomics will undoubtedly provide a much clearer picture of circRNA biology in disease. This will not only refine their use as biomarkers but will also lay a robust theoretical foundation for precision therapeutic strategies. Ultimately, circRNA-based diagnostics and therapeutics hold the promise of becoming a new, cell-free, and highly targetable modality for the clinical management of HF.
CONCLUSIONS AND FUTURE DIRECTIONS
CircRNAs represent a paradigm of multidimensional, highly integrated regulation in the molecular pathology of HF. They not only participate in myocardial remodeling by regulating key processes such as myocardial hypertrophy, cardiomyocyte apoptosis, myocardial fibrosis, inflammatory responses, and calcium signaling pathways, but also form complex signaling networks with miRNAs, RBPs, exosomes, and their own encoded small peptides, demonstrating their potential as upstream integration hubs in the HF pathology process. This bidirectional regulatory characteristic - the existence of both pro-pathogenic and protective circRNAs - provides the possibility of precise intervention, making selective molecular intervention targeting specific circRNAs or their downstream axes the theoretical basis for “directional correction” of myocardial pathological processes. Simultaneously, the stability and detectability of circRNAs in blood and EVs make them potential biomarkers for the diagnosis, risk stratification, and dynamic monitoring of HF and myocardial infarction, potentially driving the development of personalized medicine strategies.
Despite this commitment, the process of translating basic research into clinical applications is fraught with numerous challenges, including complex expression patterns, differences between animal models and human physiology, as well as significant obstacles in treatment delivery and safety. Future research will need to combine large mammalian models, human tissue samples, and exosome or nucleic acid modification technologies to enhance the feasibility and safety of circRNA-related therapeutic strategies. The key point is that current literature primarily relies on the pressure overload model, which predominantly induces concentric hypertrophy. The specific role of circRNA in diastolic hypertrophy caused by volume overload remains largely unexplored. Since pressure overload and volume overload activate distinctly different mechanical signal transduction pathways, it is imperative to conduct research using the volume overload model in the future to comprehensively characterize circRNA profiles under various biomechanical stress conditions. Moreover, given the well-recognized sex differences in HF epidemiology, remodeling patterns, and clinical outcomes, future investigations should incorporate sex as a biological variable to determine whether circRNA expression profiles and regulatory networks exhibit sex-specific characteristics[91]. Speculatively, by modulating calcium signaling and excitation-contraction coupling, circRNAs may serve as potential therapeutic targets for arrhythmias and contractile dysfunction, highlighting a novel translational avenue beyond conventional structural remodeling interventions. Furthermore, recent studies have implicated circRNAs in anthracycline-induced cardiotoxicity, in which they modulate cardiomyocyte apoptosis, oxidative stress, and mitochondrial dysfunction in doxorubicin-treated models, thereby expanding their relevance to chemotherapy-related cardiac injury[92]. In conclusion, circRNAs are not merely tools for dissecting disease mechanisms; they are foundational to a forward-thinking vision for cardiovascular medicine, representing a key translational breakthrough for diagnosing and treating HF.
DECLARATIONS
Authors’ contributions
Made substantial contributions to the conception, performed literature search, and wrote the initial draft of the manuscript: Wang X
Assisted in design of the study, literature collection, discussions, and coordination of the manuscript: Sun W
Provided critical guidance, reviewed and revised the manuscript, and approved the final version for submission: Gong W, Nie S
All authors read and approved the final manuscript.
Availability of data and materials
Not applicable.
AI and AI-assisted tools statement
During the preparation of this manuscript, the AI tools Doubao and Qwen (version Doubao-Seed-2.0, released 2026-02, and version Qwen3.5, released 2026-02) were used solely to assist in organizing the logical structure of the article and to generate a small number of supplementary icons. The tools did not influence the study design, data collection, analysis, interpretation, or the scientific content of the work. All authors take full responsibility for the accuracy, integrity, and final content of the manuscript.
Financial support and sponsorship
The study was supported by the Beijing Natural Science Foundation [24JL001] and the National Natural Science Foundation of China (82270258).
Conflicts of interest
All authors declared that there are no conflicts of interest.
Ethical approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Copyright
© The Author(s) 2026.
REFERENCES
1. Boichenko V, Noakes VM, Reilly-o’donnell B, et al. Circulating non-coding RNAs as indicators of fibrosis and heart failure severity. Cells. 2025;14:553.
2. Jones NR, Roalfe AK, Adoki I, Hobbs FR, Taylor CJ. Survival of patients with chronic heart failure in the community: a systematic review and meta-analysis. Eur J Heart Fail. 2019;21:1306-25.
4. Abbas N, Perbellini F, Thum T. Non-coding RNAs: emerging players in cardiomyocyte proliferation and cardiac regeneration. Basic Res Cardiol. 2020;115:52.
5. Zhao J, Huang H. Extracellular vesicle-derived non-coding RNAs: key mediators in remodelling heart failure. Curr Issues Mol Biol. 2024;46:9430-48.
6. Li J, Han Y, Wang S, Wu X, Cao J, Sun T. Circular RNAs: biogenesis, biological functions, and roles in myocardial infarction. Int J Mol Sci. 2023;24:4233.
7. Bibi A, Bartekova M, Gandhi S, et al. ; CardioRNA COST Action CA17129 and AtheroNET COST Action CA21153. Circular RNA regulatory role in pathological cardiac remodelling. Br J Pharmacol. 2024;182:316-39.
8. Garcia-padilla C, Lozano-velasco E, Garcia-lopez V, et al. Comparative analysis of non-coding RNA transcriptomics in heart failure. Biomedicines. 2022;10:3076.
9. Wu S, Chen L, Zhou X. Circular RNAs in the regulation of cardiac hypertrophy. Molecular Therapy Nucleic Acids. 2022;27:484-90.
10. Wei L, Liu L, Bai M, Ning X, Sun S. CircRNAs: versatile players and new targets in organ fibrosis. Cell Commun Signal. 2023;21:90.
11. Xie Q, Ma Y, Ren Z, Gu T, Jiang Z. Circular RNA: a new expectation for cardiovascular diseases. J Cell Biochem. 2023;125:e30512.
12. Uzoechina J, Zhang Z. Circular RNA-encoded proteins in disease pathogenesis. Int. J. Biol. Sci. 2025;21:6179-96.
13. Yao X, Huang X, Chen J, Lin W, Tian J. Roles of non-coding RNA in diabetic cardiomyopathy. Cardiovasc Diabetol. 2024;23:227.
14. Din ZU, Xia J, Wu Z, Liao Z, Xiong X. Circular RNAs and endothelial dysfunction: mechanistic crosstalk and emerging therapeutic perspectives. Cell Mol Life Sci. 2025;83:10.
15. Sygitowicz G, Sitkiewicz D. Involvement of circRNAs in the development of heart failure. Int J Mol Sci. 2022;23:14129.
16. Hu Y, Cao H, Sheng J, et al. Functional role of circRNA CHRC through miR-431-5p/KLF15 signaling axis in the progression of heart failure. J Genet Genomics. 2024;51:844-54.
17. Lu P, Zhang D, Ding F, Ma J, Xiang YK, Zhao M. Silencing of circCacna1c inhibits ISO-induced cardiac hypertrophy through miR-29b-2-5p/NFATc1 Axis. Cells. 2023;12:1667.
18. Xie Q, Xu X, Xiong D, Yao M, Zhou Y. CircRNA Larp4b/miR-298-5p/Mef2c regulates cardiac hypertrophy induced by angiotensin II. Int J Sports Med. 2023;45:33-40.
19. Chen C, Shen H, Huang Q, Li Q. The circular RNA CDR1as regulates the proliferation and apoptosis of human cardiomyocytes through the miR-135a/HMOX1 and miR-135b/HMOX1 axes. Genet Test Mol Biomarkers. 2020;24:537-48.
20. Wang Y, Wang H, Zhang L, Zhang J, Liu N, Zhao P. A novel identified circular RNA, circSnap47, promotes heart failure progression via regulation of miR-223-3p/MAPK axis. Mol Cell Biochem. 2022;478:459-69.
21. Han J, Zhang L, Hu L, et al. Circular RNA-expression profiling reveals a potential role of Hsa_circ_0097435 in heart failure via sponging multiple microRNAs. Front. Genet. 2020;11:212.
22. Wang Z, Liu X, Deng Y. Negative feedback of SNRK to circ-SNRK regulates cardiac function post-myocardial infarction. Cell Death Differ. 2021;29:709-21.
23. Zheng H, Shi L, Tong C, Liu Y, Hou M. circSnx12 is involved in ferroptosis during heart failure by targeting miR-224-5p. Front. Cardiovasc. Med. 2021;8:656093.
24. Li B, Bai W, Guo T, et al. Statins improve cardiac endothelial function to prevent heart failure with preserved ejection fraction through upregulating circRNA-RBCK1. Nat Commun. 2024;15:2953.
25. Huang S, Li X, Zheng H, et al. Loss of super-enhancer-regulated circRNA Nfix induces cardiac regeneration after myocardial infarction in adult mice. Circulation. 2019;139:2857-76.
26. Dong K, He X, Su H, Fulton DJR, Zhou J. Genomic analysis of circular RNAs in heart. BMC Med Genomics. 2020;13:167.
27. Li F, Du WW, Li X, et al. A new protein isoform encoded by human circular RNA circSLC8a1 contributes to cardiac remodelling. Cardiovasc Res. 2025;121:882-99.
28. Zhu Y, Zheng C, Zhang R, et al. Circ-Ddx60 contributes to the antihypertrophic memory of exercise hypertrophic preconditioning. J. Adv. Res.2023;46:113-21.
29. Li C, Wang J, Feng J, et al. Circ-TLR4 promotes cardiac hypertrophy through recruiting FUS to stabilize TLR4 mRNA. J Interv Card Electrophysiol. 2022;65:153-63.
30. Feng Y, Yan B, Cheng H, et al. Knockdown circ_0040414 inhibits inflammation, apoptosis and promotes the proliferation of cardiomyocytes via miR‐186‐5p/PTEN/AKT axis in chronic heart failure. Cell Biol Int. 2021;45:2304-15.
31. Wang LL, Yao HM, Zhao XX, Yin XW, Tian JS. Knockdown of circRNA_0030042 attenuates heart failure via miR‐568/PRG4 pathway. J Biochem Mol Toxicol. 2025;39:e70267.
32. Zhang P, Liu Y, Zhan Y, et al. Circ-0006332 stimulates cardiomyocyte pyroptosis via the miR-143/TLR2 axis to promote doxorubicin-induced cardiac damage. Epigenetics. 2024;19:2380145.
33. Gao X, Liu C, Zhang Y, et al. The circRNA CNEACR regulates necroptosis of cardiomyocytes through Foxa2 suppression. Cell Death Differ. 2021;29:527-39.
34. Deng Y, Wang J, Xie G, Zeng X, Li H. Circ-HIPK3 Strengthens the effects of adrenaline in heart failure by MiR-17-3p - ADCY6 axis. Int. J. Biol. Sci. 2019;15:2484-96.
35. Bibi A, Madè A, Greco S, et al. Circular PVT1 promotes cardiac fibroblast activation interacting with miR-30a-5p and miR-125b-5p. Cell Death Dis. 2025;16:325.
36. Guo J, Chen L, Huang Z, et al. Suppression of the inhibitory effect of circ_0036176-translated Myo9a-208 on cardiac fibroblast proliferation by miR-218-5p. J Cardiovasc Transl Res. 2022;15:548-59.
37. Ji D, Jin S, Jiang Y, et al. CircNSD1 promotes cardiac fibrosis through targeting the miR-429-3p/SULF1/Wnt/β-catenin signaling pathway. Acta Pharmacol Sin. 2024;45:2092-106.
38. Zhu Y, Pan W, Yang T, et al. Upregulation of circular RNA CircNFIB attenuates cardiac fibrosis by sponging miR-433. Front. Genet. 2019;10:564.
39. Wu N, Xu J, Du WW, et al. YAP Circular RNA, circYap, attenuates cardiac fibrosis via binding with tropomyosin-4 and gamma-actin decreasing actin polymerization. Mol Ther. 2021;29:1138-50.
40. Pang P, Si W, Wu H, et al. The circular RNA circHelz enhances cardiac fibrosis by facilitating the nuclear translocation of YAP1. Transl Res. 2023;257:30-42.
41. Ma GB, Chen WX, Zhan FJ, et al. Circ_0002295 facilitated myocardial fibrosis progression through the miR‐1287/CXCR2 axis. Clin Exp Pharmacol Physiol. 2023;50:944-53.
42. Zheng H, Huang S, Wei G, et al. CircRNA Samd4 induces cardiac repair after myocardial infarction by blocking mitochondria-derived ROS output. Mol Ther. 2022;30:3477-98.
43. Ju J, Li X, Zhao X, et al. Circular RNA FEACR inhibits ferroptosis and alleviates myocardial ischemia/reperfusion injury by interacting with NAMPT. J Biomed Sci. 2023;30:45.
44. Zang J, Lu D, Xu A. The interaction of circRNAs and RNA binding proteins: an important part of circRNA maintenance and function. J Neurosci Res. 2018;98:87-97.
45. Du WW, Yang W, Chen Y, et al. Foxo3 circular RNA promotes cardiac senescence by modulating multiple factors associated with stress and senescence responses. Eur Heart J. 2016:ehw001.
46. Xu Q, Liu J, Zhu R, et al. NSD2 promotes pressure overload-induced cardiac hypertrophy via activating circCmiss1/TfR1/ferroptosis signaling. Life Sci. 2023;328:121873.
47. Zhou L, Zhai M, Huang Y, et al. The circular RNA ACR attenuates myocardial ischemia/reperfusion injury by suppressing autophagy via modulation of the Pink1/ FAM65B pathway. Cell Death Differ. 2018;26:1299-315.
48. Lim TB, Aliwarga E, Luu TDA, et al. Targeting the highly abundant circular RNA circSlc8a1 in cardiomyocytes attenuates pressure overload induced hypertrophy. Cardiovasc Res. 2019;115:1998-2007.
49. Li M, Ding W, Fang X, et al. Novel truncated peptide derived from circCDYL exacerbates cardiac hypertrophy. Circ Res. 2025;136.
50. Wei G, Li C, Jia X, et al. Extracellular vesicle-derived CircWhsc1 promotes cardiomyocyte proliferation and heart repair by activating TRIM59/STAT3/Cyclin B2 pathway. J. Adv. Res.2023;53:199-218.
51. Liu J, Yao D, Zhou F, et al. RBM24-mediated biogenesis of circ23679 protects cardiomyocytes against apoptosis via sponging miR-15b-5p. Int J Biol Macromol. 2025;310:143242.
52. Fan X, Yang Y, Chen C, Wang Z. Pervasive translation of circular RNAs driven by short IRES-like elements. Nat Commun. 2022;13:3751.
53. Xu J, Du WW, Wu N, et al. The circular RNA circNlgnmediates doxorubicin-inducedcardiac remodeling and fibrosis. Molecular Therapy - Nucleic Acids. 2022;28:175-89.
54. Du WW, Xu J, Yang W, et al. A neuroligin isoform translated by circNlgn contributes to cardiac remodeling. Circ Res. 2021;129:568-82.
55. Legnini I, Di Timoteo G, Rossi F, et al. Circ-ZNF609 is a circular RNA that can be translated and functions in myogenesis. Mol Cell. 2017;66:22-37.e9.
56. Jiang T, Xia Y, Lv J, et al. A novel protein encoded by circMAPK1 inhibits progression of gastric cancer by suppressing activation of MAPK signaling. Mol Cancer. 2021;20:66.
57. Zhang M, Huang N, Yang X, et al. A novel protein encoded by the circular form of the SHPRH gene suppresses glioma tumorigenesis. Oncogene. 2018;37:1805-14.
58. Li Z, Huang C, Bao C, et al. Exon-intron circular RNAs regulate transcription in the nucleus. Nat Struct Mol Biol. 2015;22:256-64.
59. Qin T, Li J, Zhang K. Structure, regulation, and function of linear and circular long non-coding RNAs. Front. Genet. 2020;11:150.
60. Gasparini S, Licursi V, Presutti C, Mannironi C. The secret garden of neuronal circRNAs. Cells. 2020;9:1815.
61. Chen J, Xiao X, Zhou C, Zhang Y, Rhee J, Li H. The dark side of vascular aging: noncoding ribonucleic acids in heart failure with preserved ejection fraction. Cells. 2025;14:1269.
62. Zhang J. Non-coding RNAs and angiogenesis in cardiovascular diseases: a comprehensive review. Mol Cell Biochem. 2024;479:2921-53.
63. Yang T, Long T, Du T, Chen Y, Dong Y, Huang Z. Circle the cardiac remodeling with circRNAs. Front. Cardiovasc. Med. 2021;8:702586.
64. Wang K, Long B, Liu F, et al. A circular RNA protects the heart from pathological hypertrophy and heart failure by targeting miR-223. Eur Heart J. 2016;37:2602-11.
65. Ao X, Ding W, Li X, et al. Non-coding RNAs regulating mitochondrial function in cardiovascular diseases. J Mol Med. 2023;101:501-26.
66. Zhang Y, Chen B. Silencing circ_0062389 alleviates cardiomyocyte apoptosis in heart failure rats via modulating TGF-β1/Smad3 signaling pathway. Gene. 2021;766:145154.
67. Si X, Zheng H, Wei G, et al. circRNA Hipk3 induces cardiac regeneration after myocardial infarction in mice by binding to Notch1 and miR-133a. Molecular Therapy Nucleic Acids. 2020;21:636-55.
68. Ni H, Li W, Zhuge Y, et al. Inhibition of circHIPK3 prevents angiotensin II-induced cardiac fibrosis by sponging miR-29b-3p. Int J Cardiol. 2019;292:188-96.
69. Zhang M, Wang Z, Cheng Q, et al. Circular RNA (circRNA) CDYL induces myocardial regeneration by ceRNA after myocardial infarction. Med Sci Monit. 2020;26:923188.
70. Mckinsey TA, Foo R, Anene-nzelu CG, et al. Emerging epigenetic therapies of cardiac fibrosis and remodelling in heart failure: from basic mechanisms to early clinical development. Cardiovasc Res. 2022;118:3482-98.
71. Mei S, Ma X, Zhou L, et al. CircSMAD3 represses SMAD3 phosphorylation and ameliorates cardiac remodeling by recruiting YBX1. iScience. 2024;27:110200.
72. Jeong A, Lim Y, Kook T, et al. Circular RNA circSMAD4 regulates cardiac fibrosis by targeting miR-671-5p and FGFR2 in cardiac fibroblasts. Molecular Therapy Nucleic Acids. 2023;34:102071.
73. Ma C, Wei Z, Sun T, et al. Circ-sh3rf3/GATA-4/miR-29a regulatory axis in fibroblast-myofibroblast differentiation and myocardial fibrosis. Cell Mol Life Sci. 2023;80:50.
74. Wu N, Li C, Xu B, et al. Circular RNA mmu_circ_0005019 inhibits fibrosis of cardiac fibroblasts and reverses electrical remodeling of cardiomyocytes. BMC Cardiovasc Disord. 2021;21:308.
75. Pan W, Hunkler HJ, Chatterjee S, et al. A circular RNA derived from the ryanodine receptor 2 locus controls cardiac hypertrophy and calcium handling. Cell Mol Life Sci. 2025;82:359.
76. Zhang H, Li J, Wang Y, Li Q, Song L, Li M. CircNCX1 contributes to mitochondrial fusion in cardiomyocyte by mediating the expression of mitofusin 2. Int J Biol Macromol. 2025;329:147738.
77. Xiong J, Hu Y, Liu Y, Zeng X. CircRNA mmu_circ_0000021 regulates microvascular function via the miR-143-3p/NPY axis and intracellular calcium following ischemia/reperfusion injury. Cell Death Discov. 2022;8:315.
78. Wang K, Gan T, Li N, et al. Circular RNA mediates cardiomyocyte death via miRNA-dependent upregulation of MTP18 expression. Cell Death Differ. 2017;24:1111-20.
79. Salgado-somoza A, Zhang L, Vausort M, Devaux Y. The circular RNA MICRA for risk stratification after myocardial infarction. IJC Heart & Vasculature. 2017;17:33-6.
80. Chair S, Chan J, Waye M, Liu T, Law B, Chien W. Exploration of potential genetic biomarkers for heart failure: a systematic review. Int J Environ Res Public Health. 2021;18:5904.
81. Ibrahim NE, Januzzi JL. Established and emerging roles of biomarkers in heart failure. Circ Res. 2018;123:614-29.
82. Bayoumi AS, Aonuma T, Teoh J, Tang Y, Kim I. Circular noncoding RNAs as potential therapies and circulating biomarkers for cardiovascular diseases. Acta Pharmacol Sin. 2018;39:1100-9.
83. Sun Y, Jiang X, Lv Y, et al. Circular RNA expression profiles in plasma from patients with heart failure related to platelet activity. Biomolecules. 2020;10:187.
84. Capirossi G, Brasini S, Tremoli E, Binatti A, Roncarati R. Circular RNAs in cardiovascular physiopathology: from molecular mechanisms to therapeutic opportunities. Int J Mol Sci. 2025;26:9725.
85. Bär C, Chatterjee S, Falcão Pires I, et al. Non-coding RNAs: update on mechanisms and therapeutic targets from the ESC Working Groups of Myocardial Function and Cellular Biology of the heart. Cardiovasc Res. 2020;116:1805-19.
86. Taherpour A, Ebrahimi S, Mirzavi F. A comprehensive insight into the roles of exosomal circRNAs in metabolic syndrome. Biofactors. 2025;51:e70053.
87. Wang TZ, Thapa RK, Yu F, et al. Mining, validating, and quantifying circular RNA transcriptome from total RNA as a biomarker or target. Sci Rep. 2025;15:20418.
88. Mi Z, Zhongqiang C, Caiyun J, Yanan L, Jianhua W, Liang L. Circular RNA detection methods: a minireview. Talanta. 2022;238:123066.
89. Bakhtiarizade MR, Heidari M, Ghanatghestani AHM. Comprehensive circular RNA profiling in various sheep tissues. Sci Rep. 2024;14:26238.
90. Bahrulolum H, Rouzbahani FN, Hoseini ZS, Chaharmahali M, Hosseini SM. CircRNA: unlocking new frontiers in therapeutic and vaccine development. Mol Ther. 2026;34:748-70.
91. Chyou JY, Qin H, Butler J, Voors AA, Lam CSP. Sex-related similarities and differences in responses to heart failure therapies. Nat Rev Cardiol. 2024;21:498-516.
Cite This Article
How to Cite
Download Citation
Export Citation File:
Type of Import
Tips on Downloading Citation
Citation Manager File Format
Type of Import
Direct Import: When the Direct Import option is selected (the default state), a dialogue box will give you the option to Save or Open the downloaded citation data. Choosing Open will either launch your citation manager or give you a choice of applications with which to use the metadata. The Save option saves the file locally for later use.
Indirect Import: When the Indirect Import option is selected, the metadata is displayed and may be copied and pasted as needed.
About This Article
Special Topic
Copyright

Data & Comments
Data
0











Comments
Comments must be written in English. Spam, offensive content, impersonation, and private information will not be permitted. If any comment is reported and identified as inappropriate content by OAE staff, the comment will be removed without notice. If you have any queries or need any help, please contact us at support@oaepublish.com.